Сканирование
и форматирование: Янко Слава (Библиотека
Fort/Da) || slavaaa@yandex.ru || yanko_slava@yahoo.com || http://yanko.lib.ru || Icq# 75088656 || Библиотека: http://yanko.lib.ru/gum.html
|| Номера страниц - внизу. АНОНС КНИГИ
АНОНС КНИГИ
GENETICS
A BEGINNER'S GUIDE
B. GUTTMAN, A.
GRIFFITHS, D. SUZUKI AND T. CULLIS
ONEWORLD Oxford



БАРТОН ГУГТМАН, ЭНТОНИ ГРИФФИТС, ДЭВИД СУЗУКИ, ТАРА КУЛЛИС
ГЕНЕТИКА
Москва 2004

УДК
575
ББК
28.04 (4Вел) Г97
Гуттман
Б., Гриффитс Э., Сузуки Д., Куллис Т.
Г97
Генетика / Бартон Гуттман, Энтони Гриффитс, Дэвид Сузуки, Тара Куллис. - Пер. с
англ. О. Перфильева. - М.: ФАИР-ПРЕСС, 2004. - 448 с: ил. - (Наука & Жизнь).
ISBN 5-8183-0816-2
(рус.) ISBN 1-85168-304-6 (англ.)
Книга
излагает основные понятия генетики как науки и формирует основу для более
полного понимания современных дискуссий по поводу производства генетически
модифицированных продуктов и клонирования человека. Она поможет понять, как
работает современная наука, как она вписывается в более широкий контекст
общечеловеческой культуры, каковы ее логика и границы.
Для
широкого круга читателей.
УДК
575 ББК 28.04 (4Вел)
Все
права защищены. Никакая часть данной книги не может быть воспроизведена в какой
бы то
ни было форме без
письменного разрешения владельцев
авторских прав.
ISBN
5-8183-0816-2 (рус.) ISBN 1-85168-304-6 (англ.)
©
Guttman, Griffiths, Suzuki and Cullis 2002
Published by arrangement with One-world
Publications, Oxford, England All rights reserved
©
Издание на русском языке, перевод на русский язык, оформление. ФАИР-ПРЕСС, 2004
Электронное
оглавление. 2
ПРЕДИСЛОВИЕ.. 2
Глава 1.
ГЕНЕТИКА: ПРОШЛОЕ, НАСТОЯЩЕЕ И БУДУЩЕЕ.. 2
Поиски порядка и смысла. 2
Современный образ науки. 2
Перспективы современной генетики. 2
Глава 2. ОТ МИФА К СОВРЕМЕННОЙ НАУКЕ.. 2
Примитивный интерес к наследственности. 2
Одомашнивание растений и животных в зеркале мифа. 2
Научные теории наследственности. 2
Откуда берутся дети?. 2
Глава 3.
ЧТО ЖЕ ИМЕННО ПЕРЕДАЕТСЯ
ПО НАСЛЕДСТВУ?. 2
Строение клеток. 2
Рис. 3.1. На тонких срезах под микроскопом видно, что
сложные организмы состоят из многочисленных клеток: 2
Рис. 3.2. Строение клетки эукариотических организмов
(растений и животных) 2
Молекулярная структура. 2
Рис. 3.3. Разнообразие органических молекул, основным
элементом которых служат атомы углерода, как правило, соединенные в цепи. 2
Таблица 3.1. МОНОМЕРЫ БЕЛКОВ, АМИНОКИСЛОТЫ... 2
Рост и биосинтез. 2
Ферменты.. 2
Рис. 3.4. Структура фермента карбоксипептидазы. 2
Рис. 3.5: а - клеточная мембрана представляет собой
тонкий двойной слой молекул липидов с вкраплениями разного рода белков. 2
Синтез полимеров. 2
Клетки как фабрики по самовоспроизводству
и самообновлению.. 2
Глава 4.
РЕВОЛЮЦИОННОЕ ОТКРЫТИЕ: ЗАКОНЫ МЕНДЕЛЯ.. 2
Открытия Менделя. 2
Рис. 4.1. Объяснение результатов, полученных Менделем. 2
Родословные. 2
Другой пример: тестеры и нетестеры.. 2
Группы крови. 2
Множественные аллели и доминантность. 2
Тестовые скрещивания. 2
Вероятность. 2
Два гена и более. 2
Рис. 4.2. Решетка Пеннета, показывающая возможные
генотипы потомства от скрещивания двух индивидов, гетерозиготных по двум
независимым признакам. 2
Первый закон Менделя и определение отцовства. 2
Глава 5. ХРОМОСОМЫ,
РАЗМНОЖЕНИЕ И ПОЛ.. 2
Клетки и размножение. 2
Рис. 5.1. Микрофотография яйцеклетки, окруженной
многочисленными сперматозоидами в момент оплодотворения. 2
Митоз и клеточный цикл. 2
Рис. 5.2. Стадии митоза: профаза-хромосомы становятся
видимыми, а оболочка ядра распадается 2
Кариотип. 2
Рис. 5.3. Для составления кариотипа делящиеся клетки
распределяют на пластине 2
а. 2
б. 2
Мейоз. 2
Рис. 5.4. Процесс мейоза (в общих чертах) в клетке с двумя
парами хромосом.. 2
Рис. 5.5. При сперматогенезе из начальной клетки
образуются четыре сперматозоида, а при оогенезе только одна клетка. 2
Мейоз и законы Менделя. 2
Местонахождение генов. 2
Половые хромосомы.. 2
Нерасхождение хромосом.. 2
Рис. 5.6. Последствия нерасхождения Х-хромосом в ооците
на первом этапе мейоза и оплодотворения сперматозоидами с хромосомами X или Y. 2
Мужчины XYY: 'хромосома преступности'. 2
Попытки определения пола. 2
Глава 6.
ФУНКЦИЯ ГЕНОВ.. 2
Гены и нарушения метаболизма. 2
Рис. 6.1. Метаболические пути тирозина и фенилаланина в
организме человека. 2
Гены и ферменты.. 2
Белки и информация. 2
Рис. 6.2. Трехмерная структура белка миоглобина. 2
Рис. 6.3. Трехмерная структура гемоглобина. 2
Рис. 6.4. Электронная микрофотография красных кровяных клеток
человека, больного серповидноклеточной анемией. 2
Рис. 6.5. Последовательность первых 28 аминокислот
нормальной бета-цепи гемоглобина человека 2
Исправление наследственных нарушений. 2
Диетическое питание при фенилкетонурии. 2
Химическая коррекция серповидноклеточной анемии. 2
Перспективы эвфеники. 2
Глава 7.
НАСЛЕДСТВЕННЫЙ МАТЕРИАЛ, ДНК.. 2
Бактерии. 2
Рис. 7.1. Большая бактерия Clostridium welchii имеет приблизительные
размеры 4 х 1 мкм (микрометров). 2
Рис. 7.2. Если раствор с бактериями нанести на
поверхность питательной среды в чашке Петр 2
Первые шаги. 2
Вирусы.. 2
Бактериофаги. 2
Рис. 7.3. Общее строение вирусных частиц, или вирионов,
вирусов бактерий и вирусов животных 2
Рис. 7.4. Фаги выращивают в чашках Петри, смешивая их
образец с бактериями, в которых они могут размножаться, и с агаром. 2
Эксперимент Херши-Чейз. 2
Рис. 7.5. Схема протекания фаговой инфекции. 2
Строение ДНК.. 2
Рис. 7.6. Форма двойной спирали ДНК, по Уотсону и Крику. 2
Модель ДНК и генетика. 2
Рис. 7.7. При репликации ДНК комплекс ферментов разъединяет
цепи двойной молекулы 2
Проверка модели. 2
Глава 8.
СТРОЕНИЕ ГЕНА.. 2
Распределение генов. 2
Рис. 8.1. Генетическая карта плодовой мушки Drosophila melanogaster 2
Кроссинговер внутри генов. 2
Генетика фагов. 2
Тонкая структура гена. 2
Комплементация и определение границ гена. 2
Рис. 8.2. С помощью комплемвнтационного теста можно
определить, происходят ли две мутации внутри одного гена или нет. 2
Что же такое ген?. 2
Рестрикционные ферменты и палиндромы.. 2
Рис. 8.3. Молекулы ДНК можно легко разделить. 2
Рис. 8.4. Один из методов определения
последовательности молекулы ДНК.. 2
Рестрикционное картирование. 2
Глава 9.
РАСШИФРОВКА КОДА ЖИЗНИ.. 2
Как строятся белки! 2
Рис. 9.1. Эндоплазматическая сеть клеток эукариот состоит
из мембран, обычно расположенных параллельно друг другу и покрытых крохотными
частицами - рибосомами, которые служат фабриками по производству белка 2
Молекулы РНК: инструменты для синтеза белка. 2
РНК-транскрипция. 2
Рис. 9.2. В процессе транскрипции на одной из цепей ДНК
образуется комплементарная ей цепь РНК. 2
Рис. 9.3. Электронная микрофотография транскрипции РНК.
ДНК имеет вид тонкой нити, проходящей через центр каждого участка, похожего по
форме на перо. 2
Трансляция. 2
Рис. 9.4. Общая структура молекулы транспортной РНК. 2
Сложные гены эукариот. 2
Рис. 9.5. Общие принципы синтеза белка. 2
Генетический словарь. 2
Таблица 9.1. Генетический код. 2
Колинеарность генов и белков. 2
Рис. 9.6. Колинеарность гена и синтезируемого им белка. 2
Терминирующие кодоны.. 2
Универсальность кода. 2
Глава 10.
НАСЛЕДСТВЕННОСТЬ В МИРЕ БАКТЕРИЙ.. 2
Бактерии-мутанты.. 2
Пол у E.coli 2
Рис. 10.1. Перенос ДНК при скрещивании Hfr х F. 2
Рис. 10.2. Карта хромосомы Е. coli. 2
Факторы резистентности и устойчивость к антибиотикам.. 2
Лизогения. 2
Гены, переносимые вирусом.. 2
Трансдукция и геном человека. 2
Отбор мутантов. 2
Глава 11.
РЕГУЛЯЦИЯ ГЕНОВ И РАЗВИТИЕ ОРГАНИЗМА.. 2
Регуляция генов у бактерий. 2
Белки, которые связываются. 2
Регуляция генов эукариот. 2
Эмбриональное развитие в общих чертах. 2
Рис. 11.1. Формирование глаза эмбриона цыпленка
начинается с чашечковидного отростка мозга. 2
Регуляция по времени и развитие крыла цыпленка. 2
Рис. 11.2. Нормальное крыло птенца содержит набор костей,
обычный для всех позвоночных: 2
Регуляция по положению и развитие тела мухи. 2
Рис. 11.3. Начальные взаимодействия генов, определяющих
пространственную ориентацию эмбриона плодовой мушки. 2
Формирование глаза мухи. 2
Рис. 11.4. Восемь клеток, которые становятся
фоторецепторами в омматидии глаза дрозофилы, специализируются в определенном
порядке, который зависит от взаимодействия нескольких генов и кодируемых ими
белков 2
Глава 12.
ВМЕШАТЕЛЬСТВО В СТРОЕНИЕ ДНК: ВОЗВРАЩЕНИЕ ЭПИМЕТЕЯ?. 2
Рекомбинантная ДНК и рестриктазы.. 2
Изучение отдельных клонированных фрагментов. 2
Рис. 12.1. Для запуска полимеразной цепной реакции
отрезок ДНК нагревают до разделения его на две цепи. 2
Трансгенные организмы.. 2
Рис. 12.2. Метод ДНК-отпечатков использован для
установления невиновности одного из двух обвиняемых в изнасиловании. 2
Генная терапия. 2
Геномика - изучение всего генома. 2
Функциональная
геномика. 2
Глава 13.
ГЕНЕТИК В РОЛИ ДОКТОРА ФРАНКЕНШТЕЙНА.. 2
Контроль над исследованиями рекомбинантных ДНК.. 2
Генетически модифицированные организмы.. 2
Технологии в контексте. 2
Аргументы против генетически модифицированных продуктов. 2
Непредсказуемость генетических изменений. 2
Влияние генетически модифицированных продуктов на здоровье. 2
Возможный экологический урон. 2
Усиление власти и рост прибыли транснациональных корпораций. 2
Неестественность ДНК-технологий (ученые в роли 'богов') 2
Этические аспекты клонирования. 2
Клонирование
ДНК.. 2
Ответственность ученых. 2
Геномика и здравоохранение. 2
Глава 14.
ИСТОЧНИК ПЕРЕМЕН: МУТАЦИИ.. 2
Частота мутаций. 2
Мутации у людей. 2
Излучение. 2
Корпускулярное
излучение, или радиация, 2
Таблица 14.1 Примерные дозы излучений. 2
Что представляют собой мутации?. 2
Мутация
- это изменение в ДНК. 2
Аналоги
оснований. 2
Рис. 14.1. Дезаминирующий агент, такой как азотистая
кислота (NH02), удаляет
аминогруппу двух оснований, превращая их в основания, образующие неправильные
пары.. 2
Система восстановления ДНК.. 2
Генетические последствия радиации. 2
Хромосомные аберрации. 2
Хромосомы человека. 2
Рис. 14.2. Хромосомы человека со стандартным рисунком
Гимза-дисков. 2
Анеуплоидия. 2
Дупликация и делеция. 2
Инверсии. 2
Транслокации. 2
Глава 15.
ЭВОЛЮЦИОННАЯ ГЕНЕТИКА.. 2
Доказательства эволюции. 2
Эволюция как процесс. 2
Популяционная генетика. 2
Эволюция человека. 2
Миграция и разнообразие Homo sapiens. 2
Цвет кожи. 2
Форма тела. 2
Сопротивляемость малярии. 2
Приспособленность к высоте. 2
Евгеника. 2
ПРИМЕЧАНИЯ.. 2
Глава первая. 2
Глава вторая. 2
Глава третья. 2
Глава четвертая. 2
Глава пятая. 2
Глава шестая. 2
Глава седьмая. 2
Глава восьмая. 2
Глава десятая. 2
Глава тринадцатая. 2
Глава четырнадцатая. 2
СЛОВАРЬ.. 2
ОГЛАВЛЕНИЕ.. 2
Огромное влияние на жизнь человечества в
наши дни оказывают генетика и связанные с ней исследования физиологических
особенностей размножения человека. При написании книги мы ориентировались на
хорошо образованного читателя, который желает больше узнать об основах этой
науки, о том, какие вопросы она поставила перед человечеством, но, возможно,
немного подзабыл основы генетики в том объеме, в каком ее изучают в школе. В
книге излагаются основные понятия генетики как науки и формируются основы для
более полного понимания современных дискуссий по поводу производства
генетически модифицированных продуктов или клонирования человека.
Современные люди часто не помнят или
забывают о прошлом и считают, что интерес к генетике и связанным с ней вопросам
возник недавно. Порой даже предполагается, что споры по поводу наследственности
или искусственного воспроизводства организмов начались в связи с развитием
молекулярной генетики в последние десятилетия. Одна из задач данной книги
заключается в том, чтобы развеять это убеждение. Достаточно обратиться к
истории, почитать древние мифы и сочинения, посмотреть на самые ранние
произведения искусства наших далеких предков, дающие представление об их
чувствах и образе мысли. Можно даже утверждать, что интерес к вопросам
размножения столь же
5
древен, как и наш собственный биологический вид.
Размножение - основное проявление жизнедеятельности для любого вида, даже если
его представители этого и не осознают. Мы, конечно, никогда не узнаем, что
думали Homo habilis и
Homo
erectus миллионы
лет назад, когда они только начали познавать окружающий мир и интересоваться
вопросами существования. Однако совершенно очевидно, что в какой-то давний
период предки человека поняли, что существование их рода зависит от
размножения, то есть от рождения новых людей, и тогда они задумались, каким
образом происходит это рождение и почему дети похожи на своих родителей.
Насколько нам известно, вопросы наследственности и размножения приобрели особую
остроту, когда началось одомашнивание животных и растений.
В этой книге мы не забыли и о прозрениях
людей, вдохновленных произведениями искусства и литературы. Мы считаем, что они
должны занять достойное место в истории науки: ведь они неотделимы от
суммарного человеческого знания, поэтому старались упоминать о них там, где
необходимо. Таким образом, наша книга адресована всем, кто хочет понять, как
наука вписывается в более широкий контекст общечеловеческой культуры. Включив
примеры из искусства и истории в качестве дополнения к научному изложению, мы
постарались реалистично рассказать, как развивается современная наука. В то же
время следует осознавать логику и границы науки, рассматривая ее как часть
общего культурного феномена.
Что касается противоречий, порожденных
современной генетикой, то мы старались излагать их непредвзято и уравновешенно.
Конечно, мы не во
6
всем согласны друг с другом и потому
старались написать так, чтобы книга удовлетворила всех. Если же разногласия
оставались, то они не выходили за рамки либерального гуманизма, который, как
нам кажется, оправдан в современной биологии. Мы отнюдь не восхваляем науку во
всех ее проявлениях, поскольку осознаем опасность практически каждого открытия;
вместе с тем мы и не 'луддиты', критикующие любые научные и технические
достижения. Когда была разработана технология рекомбинантных ДНК, многие
уважаемые ученые предупреждали о возможных опасностях этой технологии. Конечно,
специалисты обязаны были предвидеть все опасности и предупредить о них, но
теперь, когда разработаны разумные методы контроля, совершенно очевидно, что
благодаря новым технологиям человечество может получить много преимуществ и
благ с минимальными негативными последствиями. Ко всякому открытию следует
подходить с такой же ответственностью. Каждое открытие ставит серьезные
социальные и моральные вопросы, которые образованные и информированные члены
общества обязательно должны обсуждать. Мы постарались дать хотя бы краткое
изложение этих вопросов, чтобы читатели узнали об основных точках зрения.
Мы верим также в человечество как вид и в
биологическую равноценность всех людей, без исключения. В эпоху, когда многих
людей оскорбляют только за то, что у них 'не тот' цвет кожи или что они говорят
'не на том' языке, когда во многих странах активизируется движение неонацистов,
ученые должны понять, что наука не может оставаться 'морально нейтральной', как
выразился ученый и писатель Ч. П. Сноу, и поэтому должны четко излагать
7
основные понятия биологии. Среди групп
людей наблюдается большое генетическое разнообразие, но все отличия
укладываются в общий диапазон признаков человека и не доказывают врожденного
превосходства или неполноценности той или иной группы. Мы постарались хотя бы
вкратце затронуть нравственные вопросы, обращаясь к тем, кто еще не определил
своего отношения к ним.
Глава 1. ГЕНЕТИКА: ПРОШЛОЕ, НАСТОЯЩЕЕ И БУДУЩЕЕ
'Почему у Джимми рыжие волосы, как у
мамы, а у его папы черные?'
'Почему у людей не рождаются щенки?'
'А если конь женится на корове, у них
будут дети?'
'Почему Мэри такая высокая, а ее родители
низкорослые?'
Детские вопросы, такие наивные и
простодушные, зачастую проникают в самую суть таинственных загадок жизни.
Подобные вопросы всегда волновали философов и ученых, начиная с глубокой
древности. В поисках ответов люди обращались к мифам, суевериям и так
называемому здравому смыслу.
Известно, что все без исключения живые
организмы продолжают свой род поколение за поколением: у коров рождаются
телята, семена моркови вырастают в зрелую морковь, женщины рожают детей.
Авторов Библии этот неизменный закон поражал настолько, что они сочли достойным
упомянуть его на страницах этой великой книги и объясняли его божественным
вмешательством (Быт. 1:11, 21):
9
И сказал Бог: да произрастит земля
зелень, траву, сеющую семя, дерево плодовитое, приносящее по роду своему плод,
в котором семя его на земле.
И сотворил Бог рыб больших и всякую душу
животных пресмыкающихся, которых произвела вода, по роду их, и всякую птицу
пернатую по роду ее.
Вместе с тем отдельные представители вида
часто весьма сильно отличаются друг от друга по внешнему облику и форме.
Достаточно посмотреть на людей, проходящих по любой городской улице. Каждый
ребенок обязательно наследует черты своих родителей. Словно внутри человека
находится предписание производить потомство не только 'по роду своему', но и по
своему росту, весу, цвету кожи, глаз, волос и т. д. Люди всегда считали это
чем-то само собой разумеющимся, но не могли найти подходящего объяснения.
Возникали различные мифы и суеверия.
Порой люди настолько отличаются друг от
друга, что возникает вопрос: 'А не может ли у женщины родиться нечто, вовсе не
похожее на человека?' И в самом деле, иногда рождаются дети с очень большими
аномалиями развития, но эти случаи очень редки, и человеческая фантазия
преобразовала их в фантастические существа. Обычно у людей рождаются самые обычные
дети, правда, с уникальными признаками, делающими их неповторимыми среди
окружающих представителей нашего вида. Так откуда же такое постоянство и
одновременно различие? Этот биологический парадокс удалось разрешить только
благодаря недавним открытиям в исследовании законов наследственности. Наука,
изучающая наследственность, называется генетикой.
10
Как наука генетика возникла в 1900-х
годах, когда были сформулированы основные законы передачи наследственных
признаков от одного поколения к другому. Эти законы, применимые ко всем
растениям, животным и даже к одноклеточным организмам, доказали фундаментальную
общность всего живого и позволили людям вмешиваться в процессы
наследственности, конструировать свои формы живого. Генетики научились
создавать высокопродуктивные породы домашних животных, сорта растений и штаммы
грибков, производящих антибиотики, не говоря уже об экзотических цветах и
красивых золотых рыбках. По мере того как мы постигаем молекулярные основы
жизни, все наши мечты о создании новых ее форм переходят из области научной
фантастики в реальную жизнь. Телевидение и газеты почти ежедневно сообщают о
достижениях в области генной инженерии.
Применив полученные знания в медицине, мы
узнали, как передаются наследственные заболевания, как наследуются те или иные
черты внешности или особенности поведения. Эти открытия обнажают суть
человеческой природы. Генетика наряду с эндокринологией, физиологией и
эмбриологией многое дала для понимания человека как вида, но в то же время
поставила очень серьезные нравственные и этические вопросы. Например, в каком
случае родителям стоит решиться на прерывание беременности: в случае
значительного физического или психического дефекта, заячьей губы или просто
нежелательного пола ребенка? Когда развивающийся плод становится человеком в
полном смысле этого слова? (И стоит ли вообще задавать такой вопрос?) В 1947
году, понимая, как велика мощь незадолго
11
26
искусстве, на самом деле враги науки. В
этих популярных образах массовой культуры отразился наш оправданный страх перед
обезличенным, чистым знанием, лишенным этической составляющей, страх перед тем,
что ученые, эти достойные люди, обернутся титанами, создающими чудовищных
монстров.
Наука не может быть оторвана от общества;
именно культура ставит перед наукой определенные задачи и вопросы, а наука, в
свою очередь, оказывает влияние на культуру. Мы считаем, что только
образованное общество сможет уравновесить предоставляемые наукой силу, власть и
выгоду заботой об общественном благе. Цель данной книги - поместить науку о
наследственности в социально-исторический контекст, дать полное представление о
ее месте в современном обществе и о возможных последствиях развития генетики.
Обратимся к первым проблескам
человеческого сознания и посмотрим, как люди в древности пытались найти ответы
на интересующие их вопросы размножения, производства потомства и
наследственности.
Глава 2. ОТ МИФА К СОВРЕМЕННОЙ НАУКЕ
Генетика своими корнями уходит в далекое
прошлое, и она вовсе не возникла неожиданно, на пустом месте, в начале ХХ века.
Люди начали задумываться о наследственности, пожалуй, еще с первыми проблесками
сознания. Цивилизация многим обязана тому, что люди научились успешно разводить
домашних животных и выращивать культурные растения.
В течение долгого времени наши
первобытные предки жили подобно другим представителям царства животных, то есть
занимались охотой и собирательством, добывая себе пищу. Но в ходе эволюции у
людей развился большой и сложный мозг, который позволял находить закономерности
в окружающем мире. Этот мозг позволяет нам запоминать, учиться на примере
других, не повторять известных ошибок и открывать новое. Только около 10 тысяч
лет назад некоторые люди эпохи неолита - скорее всего, женщины, которые следили
за стоянкой, пока мужчины охотились, - пришли к мысли, что можно самим
выращивать полезные растения. Во многих местах, таких как Иерихон в долине
Иордана, или на ежегодно затопляемых равнинах Египта, люди сажали семена
растений во влажную почву, они вырастали и давали еще боль-
28
ше семян. Имея надежный запас пищи, можно
было не кочевать с места на место, охотясь на животных и собирая растения, а
жить оседло и заниматься сельским хозяйством. Так из кочевников и охотников
люди превратились в земледельцев.
Поначалу земледельцы отбирали семена
лучших растений и выращивали их неосознанно. Они поедали съедобные злаки и
овощи, а случайно упавшие на землю семена вырастали в новые растения. Дикие
животные (собаки, козы, коровы и овцы) подходили к жилищам людей, привлекаемые
объедками; некоторых из них люди отлавливали и содержали в загонах, чтобы иметь
запас мяса, шкуры и дополнительную силу для перевозки тяжестей. Расчищая
большие участки земли, земледельцы сажали фруктовые деревья, создавая первые
сады. Так Homo sapiens поднялся
на очередную ступень в своем развитии.
По всей видимости, переход к земледелию и
скотоводству время от времени предпринимался по всему земному шару и во многих
случаях закончился провалом. Окончательно земледелие утвердилось в двух
регионах: в Азии - от Междуречья до Китая около 9000-7000 лет до н. э. и в
Америке - от Мексики до Перу около 5000-2000 лет до н. э. При этом не следует
недооценивать развитие сельского хозяйства в Северной Америке: ведь около 60%
современных культурных растений было неизвестно до плаваний Колумба. Список
разводимых в наше время животных и растений представляет собой лишь малую часть
того, что пытались разводить люди за всю историю человечества.
Со временем люди поняли, что растения и
животные дают потомство 'по роду своему', то есть из
29
семян растений с большими плодами,
вероятнее всего, вырастают растения с большими плодами, а у овец с тонкой
шерстью, вероятнее всего, будет потомство с такой же тонкой шерстью. Как только
люди усвоили правило, согласно которому 'подобное порождает подобное', они
получили новый инструмент воздействия на природу.
Земледелие и скотоводство оказали
поистине грандиозное влияние на развитие человечества. Этот интеллектуальный
прорыв революционным образом заменил эволюцию биологическую эволюцией
культурной, ставшей основной движущей силой истории. Домашние растения и
животные помогли создать более стабильное общество, поскольку люди больше не
зависели от ненадежных охоты и собирательства. Население постепенно росло,
площадь расчищенных земель расширялась, и люди стали специализироваться в том
или ином виде деятельности, удовлетворяя постоянно растущие потребности
общества. Необходимо было строить защитные сооружения от набегов варваров,
делать орудия труда для обработки земли и сбора урожая, хранить урожай и
доставлять в поселок строительные материалы со все больших расстояний.
Разделение труда высвободило время для размышлений, фантазии и изобретений,
которые ускорили культурное развитие. Люди научились делать горшки, ткать
материю для одежды, плавить металлы; они усилили свою власть над природой и над
собственной судьбой. Таким образом, цивилизация возникла, когда кочевые
охотники и собиратели превратились в земледельцев, разводящих растения и
животных.
30
Примитивный интерес к наследственности
Если заглянуть в прошлое, то
свидетельства интереса к вопросам наследственности можно найти еще в период
палеолита, когда люди только начали понимать, что такое размножение. Возьмем
для примера рисунки совокупляющихся и рожающих животных и людей на стенах
пещер, выполнявшие двоякую роль. Считалось, что они посредством симпатической
магии способствуют увеличению потомства людей и животных, на которых люди
охотились; в то же время подрастающее поколение получало представление о
жизненных циклах и функциях живых организмов. Эти рисунки свидетельствуют о том,
что первобытные люди уже понимали некоторые генетические закономерности,
которые находили отражение в легендах и мифах. К тому времени, когда были
созданы циклы мифов, люди прекрасно знали, что можно улучшать свойства растений
и животных, если подбирать родителей с нужными признаками. Некоторые мифы
свидетельствуют о растущем интересе к законам наследственности и служат важными
вехами в истории генетики.
Для мифов характерно правило, согласно
которому то, что происходит среди богов, является отражением того, что
происходит на земле... Таким образом, миф представляет собой как бы
стенографическую запись вторжений, миграций, смен династий, распространения
иноземных культов и перемен в обществе. Когда в Грецию впервые попал хлеб - до
того там были распространены только бобы, семена мака, желуди и корни асфоделя,
- его употребление освятил миф о Деметре и Триптолеме; то же самое отразилось и
в валлийском мифе о Белой Старухе, богине посевов,
31
которая ходила по всей стране, сея зерно
и разводя пчел; ведь о земледелии, разведении свиней и пчеловодстве местные
жители узнали от той же волны переселенцев эпохи неолита. Другие мифы освящали
употребление вина1.
Всеобщее распространение подобных мифов о
выращивании растений и одомашнивании животных говорит о том, что интерес к
проблемам размножения и воспроизведения потомства был присущ всем древним
культурам и цивилизациям. Усвоив принципы земледелия и искусственного отбора,
человечество заинтересовалось тем, как происходит размножение у людей.
Разнообразные мифы предлагали любопытные объяснения того, откуда берутся дети и
от чего зависит пол ребенка. Далее мы рассмотрим некоторые из этих первых
объяснений.
Одомашнивание растений и животных
в зеркале мифа
В многочисленных рисунках, произведениях
изобразительного искусства и мифах древние люди отразили появление каждого из
культурных растений и одомашненных животных, оказавших очень важное влияние на
развитие человеческого общества. Часто для каждой породы животных или сорта
растений люди придумывали своего бога-покровителя. Поклоняясь этим богам,
принося им жертвы и показывая, насколько они ценят их дары, люди пытались
обрести власть над капризной природой и обеспечить свое благополучие.
Боги-защитники растений или животных олицетворяют собой то глу-
32
38
де Аристотеля, которые стали отделять
вымысел от фактов, вследствие чего наука все более удалялась от суеверий,
фантазии и заблуждений. Первые объяснения механизма наследственности нельзя
назвать научными, но они по крайней мере говорили о том, что такой механизм
существует и что человеку, при должном старании, вполне по силам его понять.
Научные теории наследственности
Сходство детей и их родителей отмечается
всеми. Древние люди считали, что похожие люди имеют общих предков, и поэтому
особое внимание уделяли родству. Помимо того что родственные связи скрепляли
древнее общество, они породили представление о том, что наследственные признаки
передаются 'по крови', что существует так называемое кровное родство. В мифах и
легендах отражено убеждение древних в том, что черты характера и поведения
определяются наследственностью; длинные перечисления в Ветхом Завете, кто кого
'родил', призваны утвердить положение человека в мире посредством его связей со
своими предками. Влиянию окружения и внешней среды придавалось меньшее
значение; предполагалось, что незнакомые люди, имеющие общих предков, гораздо
ближе друг другу, чем проживающие по соседству, но не связанные узами родства.
Индейцы Северной Америки среди всех ритуалов превыше всего почитали обычай
кровного братства, когда неродные друг другу люди смешивали свою кровь.
Родство всегда предоставляло обществу
возможность не только социального устройства, но и
39
43
Откуда берутся дети?
Огромная роль наследственности для
общества не только с физиологической, но и с культурной точек зрения, а также
заинтересованность в здоровом потомстве заставили человечество задуматься,
каким же образом происходит размножение. Отсюда оставался только один шаг к
идее контроля над этим процессом. Зная, как размножаются растения, древние люди
предположили, что мужчина помещает внутрь женщины свое 'семя'. Древние египтяне,
например, понимали этот процесс лишь частично, для них основная его часть
оставалась скорее чудом, нежели биологическим явлением. Это чудо они
приписывали богу солнца Ра, который якобы сам (но тем не менее в обличье
фараона) оплодотворял жену фараона и порождал царских преемников, которые таким
образом обретали божественное происхождение. Египетский царь Ахенатон (Эхнатон,
1379- 1351 до н. э.) воспел могущество бога в своем 'Гимне солнцу':
Ты тот, кто порождает ребенка в женщине,
Кто производит семя в мужчине,
Кто дает жизнь сыну в теле его матери,
Кто утешает его, чтобы он не плакал,
Лелеет его еще во чреве,
Кто дарит дыхание, оживляющее все,
что он ни сотворит.
По всей видимости, более или менее
последовательное учение о размножении разработали древние греки. К тому
времени, когда Гомер написал 'Илиаду', греки уже понимали, что физические
признаки и черты характера передаются по наслед-
44
50
На протяжении XVIII века, когда искали порядок, смысл и законы во всем
окружающем мире, на весьма интересный вопрос 'откуда берутся дети?' обращали
внимание десятки ученых. Так называемые 'сперматисты' придерживались теории
Левенгука и верили в то, что в каждом сперматозоиде содержится отдельный
индивид (гомункул).
Итальянец Марчелло Мальпиги с этим не
согласился; он стал родоначальником школы 'овистов', предположив, что крохотный
гомункул содержится в женском яйце (существование которого оставалось
гипотетическим) и просто 'пробуждается', когда в яйцо проникает сперматозоид.
Соперничество между этими двумя школами мысли продолжалось на протяжении десятилетий.
Француз Мопертюи высказал предположение,
что семенная жидкость каждого из родителей содержит 'частицы', ответственные за
тот или иной признак, и что эти частицы смешиваются между собой, прежде чем
образовать зародыш. Такой зародыш походит сразу на обоих родителей. Избыток
частиц порождает уродов (как нам сейчас известно, дети с синдромом Дауна имеют
лишний генетический материал), впрочем, как и недостаток частиц. Далее Мопертюи
предположил, что неиспользованные частицы остаются в организме и могут
проявляться в последующих поколениях, что объясняет возможное сходство детей с
дедушками или прадедушками.
Эта теория уже больше походит на
генетическую теорию ХХ века, нежели на древние теории Гиппократа и Аристотеля.
И наконец, в 1827 году Карл Эрнст фон Бэр открыл яйцеклетку у млекопитающих,
проложив тем самым дорогу современной генетике.
Глава 3. ЧТО ЖЕ ИМЕННО ПЕРЕДАЕТСЯ
ПО НАСЛЕДСТВУ?
Что скрывается за высказыванием 'Организм
наследует те или иные признаки'? Возьмем для примера семью, в которой у матери
ярко-рыжие волосы и зеленые глаза, тогда как у отца черные волосы и карие
глаза. У одного из их сыновей ярко-рыжие волосы, у другого рыжевато-коричневые,
а глаза у них карие или светловато-коричневые. У обоих родителей мочки ушей
выделяются отчетливо, но у одного из сыновей мочки ушей плотно прижаты к щекам.
И мать, и отец выше среднего роста, их дети также довольно высокие для своего
возраста. Дети, очевидно, унаследовали признаки от своих родителей, хотя иногда
встречаются и необычные, например форма мочек ушей. Но перед тем как начать
рассуждать о наследственности, нужно выяснить, что же такое, собственно,
признаки.
Цвет во всех растительных и животных
организмах определяют химические вещества - пигменты.
Пигмент поглощает определенную часть спектра и отражает другую его
часть; мы воспринимаем эти участки спектра отраженного света как цвета, тогда
как весь спектр кажется нам белым. Черная, бурая или рыжая окраска всегда
зависит от пигментов.
52
53
ка стремится узнать, как факторы
наследственности, называемые генами, определяют
производство определенных пигментов, гормонов или тканей. Понять это можно,
только ознакомившись с основными биологическими структурами.
Как телескоп революционным образом
преобразил астрономию, так и микроскоп помог людям понять, из чего состоят
живые организмы. Можно представить, какое удивление и изумление отразилось на
лицах ученых, когда они впервые увидели мельчайшие живые существа в капле воды
или в кусочке почвы. Левенгук оставил описания 'миниатюрных зверьков',
содержащихся в сперме и крови, которые казались ему живыми существами. В 1665
году Роберт Гук исследовал с помощью микроскопа тонкий срез пробки, через
который проходил свет, и заметил повторяющиеся ряды мелких клеточек. Он так и
назвал их - клетки, хотя на самом
деле это были лишь оболочки давно умерших клеток. Вскоре и другие исследователи
обнаружили, что все растения и животные состоят из похожих 'кирпичиков' самых
разных форм и размеров. Позже оказалось, что эти клетки в свою очередь содержат
в себе еще более мелкие структуры, необходимые для их жизнедеятельности.
В 1839 году ботаник Маттиас Якоб Шлейден
и зоолог Теодор Шванн высказали гипотезу об универсальности клеточного
строения. Они пришли к мнению, что все живые организмы развиваются из
одной-единственной клетки и что многоклеточные
54
организмы образуются в результате деления
клеток. Одно из важнейших положений современной биологии гласит, что все
организмы представляют собой либо одну клетку, либо комплекс клеток и что
клетка - это основная биологическая единица, окруженная мембраной, которая
отделяет ее внутреннюю среду от внешней среды. Таким образом, клетка - это
мельчайшая единица живой материи, и все живое, по
определению, обязательно состоит из клеток.
На рис. 3.1 в увеличении показаны два
поперечных среза - срез небольшого червя и стебля растения. Легко заметить, что
обе структуры состоят из маленьких 'кирпичиков'-клеток, плотно прижатых друг к
другу. В одном и том же организме бывают разные клетки, которые образуют разные
виды тканей, такие как эпидермис (кожа
или поверхностный слой), мышцы животных или древесина растений. Каждая ткань
состоит из особого типа клеток, но для всех клеток характерно наличие оболочки,
Рис. 3.1. На тонких срезах под
микроскопом видно, что сложные организмы состоят из многочисленных клеток:
а - небольшой червь; б -
стебель растения
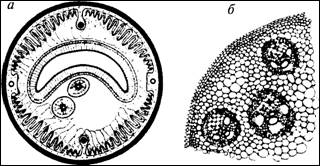
55
56
торые называются органеллами (рис. 3.2). Самой заметной
органеллой часто бывает ядро - центральная структура, ограниченная мембраной.
Ядро особенно важно для генетики, так как в нем имеются хромосомы, содержащие наследственный материал.
В клетке есть также многочисленные вытянутые тельца, которые называются митохондрии; они получают энергию из молекул
пищи, таких как сахар (глюкоза), и накапливают их в химической форме, после
чего клетка уже может использовать эту энергию. Во многих растительных клетках
имеются ярко-зеленые хлоропласты, усваивающие
энергию солнечного света и накапливающие ее также в химической форме. Обе эти
разновидности органелл состоят в основном из мембран, то есть тонких пла-
Рис. 3.2. Строение клетки
эукариотических организмов (растений и животных)
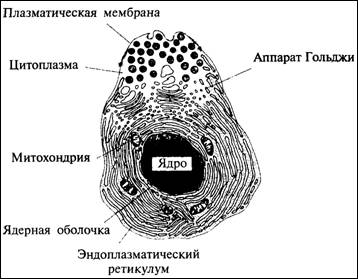
57
58
Все составные части клеток, которые можно
видеть под микроскопом, состоят из более мелких химических структур, от которых
зависит природа наследственности. Чтобы получить некоторое представление об
этих структурах, нам потребуются только самые общие сведения из курса химии, а
именно то, что все вещества состоят из атомов, которые объединяются в молекулы,
и что формула вещества отражает состав его молекул. Например, формула воды - Н2O, поскольку каждая ее молекула состоит из двух атомов
водорода (Н), связанных с одним атомом кислорода (О). Следует также вспомнить,
что атомы каждого элемента имеют определенную массу. Масса атома водорода равна
единице, углерода - 12 единицам, а железа - 55,85. Масса молекулы равна сумме
масс, составляющих эту молекулу атомов.
Рассмотрим для начала два объекта - алмаз
и кальцит, структура которых довольно характерна для обычного вещества:
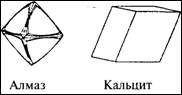
В природе часто встречаются вещества
подобного рода. Мы видим, что они имеют упорядоченную форму, и этому есть свои
причины, что станет ясно
59
при делении вещества на все более мелкие
части. Отложим в сторону алмаз (наш бюджет не позволит проводить с ним
эксперименты) и начнем дробить кальцит при помощи долота и молотка. Он
распадется на мелкие куски, но - что самое интересное - эти куски будут
повторять структуру большого куска. Не обращая внимания на размеры, можно
заметить, что углы между гранями и плоскостями остаются постоянными. Раздробив
минерал на мельчайшие частички и рассмотрев их под микроскопом, мы увидим все
ту же, уже известную нам форму. Оказывается, такое строение имеют даже
мельчайшие частички вещества.
Химики, которые называют кальцит
карбонатом кальция, скажут, что его структура состоит из карбонатной группы (СO3,
в которой атом углерода соединен с тремя атомами кислорода) и одного атома
кальция. Физические наблюдения показывают, что многочисленные карбонатные
группы и атомы кальция расположены в пространстве под теми же углами, что и
грани большого кристалла кальцита.
Таким образом, видимая структура
материала повторяет кристаллическую структуру. Это та же структура, только во
много раз увеличенная.
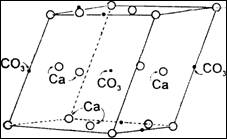
60
61
ненных между собой химическими связями. При химической связи два
атома делят между собой пару электронов - по одному от каждого атома. На наших
рисунках связь между двумя атомами изображена в виде линии. Каждый элемент
характеризуется валентностью, или
способностью образовывать определенное число химических связей. Валентность
углерода равна четырем, поэтому каждый атом углерода может быть связан с
четырьмя другими атомами; благодаря этому его свойству образуется большое число
самых разных сочетаний атомов, что приводит к огромному разнообразию
органических молекул (рис. 3.3). Две и три параллельные линии означают двойную
и тройную связь соответственно. Связь посредством пары электронов называется ковалентной; она очень прочная, для ее разрыва
требуется значительное количество энергии, потому органические молекулы
довольно стабильны. Однако связи легко разрываются при сгорании (окислении),
высвобождая большое количество энергии, поэтому углеводороды служат ценным
видом топлива.
В самой простой органической молекуле
метана атом углерода связан только с четырьмя атомами водорода. В другой
молекуле атом углерода соединен одной связью с другим атомом углерода, образуя
цепь С-С, на концах которой располагаются атомы водорода. Цепь С-С может
достигать очень большой длины; молекулы воска, например, состоят из 30-36
атомов углерода. Цепь атомов углерода может также замыкаться в кольца
различного размера. Но самое большое разнообразие получается от соединения
атомов углерода с группами атомов других элементов. Например, гидроксильная
группа ОН
62
Рис. 3.3. Разнообразие
органических молекул, основным элементом которых служат атомы углерода, как
правило, соединенные в цепи.
Каждая линия между
атомами соответствует связи, то есть общей паре электронов. Двойные и тройные
линии обозначают двойные и тройные связи между атомами. Более сложные молекулы,
особенно те, что имеют кольцевые структуры, обычно изображаются в виде линий, в
местах соединения которых атомы углерода (часто с
одним или двумя атомами водорода) не
обозначаются. Поскольку валентность углерода равна четырем, каждый атом
углерода должен иметь четыре связи; если показаны только три связи атома
углерода, то с этим атомом должен быть связан еще один атом водорода
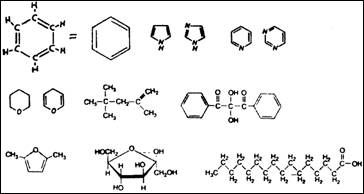
(кислород, связанный с водородом),
присоединенная к углеродной цепи, образует спирт
(алкоголь). Аминогруппа,
состоящая из атома азота и двух атомов водорода (NH2),
соединенная с углеродной цепью, образует амин. В
более сложных группах атом кислорода связан с атомом углерода двойной связью (С=0),
и одна из таких комбинаций, карбоксильная группа
COOH, образует молекулу кислоты. (Кислотой называется любое
химическое соединение,
63
69
♦ белки переносят ионы и небольшие
молекулы через клеточные мембраны, что необходимо для работы нашей нервной
системы и таких органов, как почки;
♦ белки регулируют все виды
процессов и следят за тем, чтобы они происходили с нужной скоростью.
Понять, каким образом устроены клетки и
как они работают, можно, только узнав подробнее о некоторых функциях белков.
Рост и биосинтез
Одно из самых очевидных свойств живого
организма - способность к росту. Рост любого организма, например человека,
является результатом двух процессов: роста клеток
и их деления. Человек, как и многие
другие виды организмов, растет только до известного предела, по достижении
которого размеры остаются более или менее неизменными. Однако все наши ткани
постоянно обновляются, некоторые даже с очень большой скоростью; отдельные
клетки организма также постоянно растут, на смену старым приходят новые.
Очевидно, что организм растет за счет поступления питательных веществ, из
которых он создает свои структуры. Как давно было известно, 'человек есть то,
что он ест'. Часть молекул, поступающих в виде пищи, мы преобразуем в
строительные вещества для наших клеток, а другие молекулы образуют отходы,
такие как углекислый газ, вода и мочевина. Фотосинтезирующие организмы (зеленые
растения) в свою очередь этими 'отходами' питаются, производя из них
70
72
ществ для получения энергии, которая
затем используется в биосинтезе и других процессах, но мы пока не будем
затрагивать эту тему.) Есть свой путь для каждой из 20 аминокислот, из которых
состоят белки; для каждого сахара, из которых состоят полисахариды; для каждого
липида и т. д. Все конечные продукты этих путей затем встраиваются в такие
сложные структуры, как белки или клеточные мембраны.
Линию сборки на заводах обслуживают люди (хотя
теперь их все чаще заменяют роботы). Кто же обслуживает пути метаболизма в
организме? Каким образом происходят химические реакции, превращающие один
метаболит в другой? Иногда для этого ничего дополнительного не требуется.
Некоторые химические реакции происходят быстро и сами по себе, едва только
необходимые вещества смешиваются между собой; для этого достаточно той энергии,
с которой молекулы сталкиваются. Но организмы не могут полагаться на реакции,
которые происходят сами по себе. Многие реакции просто не начнутся без
дополнительной энергии, и клетки выработали механизм подачи энергии для таких
реакций. Кроме того, многие реакции метаболизма происходят настолько медленно,
что их обязательно нужно ускорять. Для этого и существуют ферменты. Фермент - это белок,
взаимодействующий с некоторой молекулой - субстратом,
заставляющий молекулу вступать в определенную химическую реакцию.
Фермент может по очереди воздей-
73
74
только потому, что ферменты и субстраты
перемешаны внутри небольшого пространства клетки.
На рис. 3.4 показана структура одного из
ферментов, в котором цепь аминокислот, образующая его первичную структуру,
особым образом сложена и образует активный центр. На этом участке определенные
остатки аминокислот расположены так,
Рис. 3.4. Структура фермента
карбоксипептидазы.
На рисунке показано, как
аминокислоты (обозначенные буквенными сокращениями с порядковыми
номерами) образуют активный центр
именно той формы, которая необходима для проведения определенной химической
реакции. Этот пищеварительный фермент расщепляет белковые молекулы пищи. В
реакции участвует ион цинка (Zn). Поперечными черточками обозначены взаимодействия атомов
и молекул
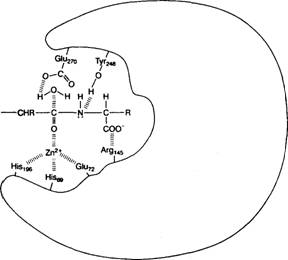
75
77
виды молекул внутри клетки и выводить
другие наружу, регулируя таким образом состав внутриклеточной жидкости.
При первичных метаболических процессах
синтезируются все аминокислоты, сахара, липиды и другие небольшие молекулы
клетки, которые идут на образование таких макромолекул, как белки и
полисахариды. После этого синтезировать простой полимер вроде целлюлозы не так
уж сложно. Молекула целлюлозы состоит из множества молекул глюкозы, и один из
ферментов соединяет их в одну большую цепь.
Синтез белков более сложный. Белки
состоят из 20 аминокислот, которые могут соединяться в любом количестве и в
любом порядке. Для каждого белка характерна определенная последовательность
аминокислот. Любая клетка, например костного мозга, синтезирующая гемоглобин,
должна получить 'инструкции', в какой последовательности следует соединять
аминокислоты. Другими словами, ей необходима информация.
Именно благодаря информации мы делаем правильный выбор из множества
возможных вариантов. Если нужно набрать телефонный номер друга или узнать
высоту Эйфелевой башни, необходимо узнать верное число из всех возможных чисел.
Точно так же информация о структуре белка определяет последовательность его
аминокислот, например: Ser-Gly-Ala-Ala-Val- Glu-His-Val-... и т. д. Отсюда следует вывод, что в организме
должны быть какие-то носители инфор-
78
79
мов. Это положение и составляет основу
современного учения о наследственности.
Клетки как фабрики по самовоспроизводству
и самообновлению
Постараемся еще раз представить, как
работает организм. Из окружающей среды он получает вещество-сырье и по
различным путям метаболизма превращает его в молекулы своей структуры - делает
из первичных продуктов мономеры, а затем и полимеры. Но что представляют собой
полимеры? В большинстве случаев это те же ферменты, с помощью которых организм
производит мономеры, а затем и полимеры. Идея ясна: организм состоит из
структур, которые производят сами себя. Белки получают информацию о своем
производстве от молекул нуклеиновых кислот, в основном от
дезоксирибонуклеиновой кислоты, или ДНК. Полный набор ДНК образует геном клетки. В то же время геном - это
физическая структура, содержащая все гены. Наследственная, или генетическая, информация определяет, как
производить структуры жизни - катализаторы, да и сам геном. Как только клетка
накапливает достаточно новых веществ и структур, она делится, после чего
процесс начинаются заново. Поэтому можно сказать, что клетка, эта
фундаментальная единица жизни, представляет собой не что иное, как механизм,
запрограммированный на самовоспроизводство.
Настало время рассмотреть подробности
этого процесса.
Глава 4. РЕВОЛЮЦИОННОЕ ОТКРЫТИЕ: ЗАКОНЫ МЕНДЕЛЯ
Тайна передачи признаков по наследству
всегда привлекала людей. В I веке до н. э.
древнеримский философ Лукреций заметил, что дети иногда походят на своих
дедушек или прадедушек. Столетием спустя Плиний Старший писал: 'Часто бывает
так, что у здоровых родителей рождаются дети-калеки, а у родителей-калек -
здоровые дети или дети с тем же уродством, в зависимости от случая'. Еще первые
земледельцы поняли, что некоторые признаки зерновых, например пшеницы, или
домашних животных, например овец, передаются по наследству, поэтому путем
отбора можно создавать новые сорта растений и новые породы животных. И сейчас
мы порой восхищенно восклицаем: 'У него улыбка матери!' или 'У нее характер
отца!', особенно когда хотим сказать, что эти черты достались детям по
наследству от родителей1.
Несмотря на то что о наследственности
люди знали давно, природа этого механизма оставалась для них скрытой.
Невозможно было дать понятное объяснение наследственности или определить законы
передачи тех или иных признаков. Самое первое и простое, приходящее на ум, -
предположение, что
81
признаки родителей в равной степени
'перемешиваются' в детях, потому дети должны представлять собой нечто среднее. Это все равно, что взять банки с
красной и белой красками, перемешать их и получить розовый цвет. Отсюда может
возникнуть предположение, что не только простые признаки (цвет волос и глаз или
форма носа), но и сложные, вроде манеры поведения или черт характера, будут
отражать нечто среднее между признаками родителей. Однако при дальнейшем
смешении красок никогда не получится чистый цвет; из розовой краски нельзя получить
чисто белую или чисто красную. Уже древние римляне около 2 тысяч лет назад
понимали, что наследственные признаки передаются каким-то другим способом.
Кроме того, на такие сложные признаки, как характер или умственные способности,
огромное влияние оказывает внешняя среда, в частности воспитание.
В теории эволюции Чарльза Дарвина
передача признаков по наследству была ключевым элементом. Скотоводы выводят
новые породы домашних животных методом искусственного отбора, природа же
создает новые виды посредством естественного отбора. Если бы полезные для
выживания признаки не наследовались, не было бы эволюции. Однако при объяснении
механизма наследственности Дарвин придерживался ошибочной теории пангенезиса.
Как мы уже видели, согласно этой гипотезе, каждый орган производит крохотные
элементы (пангены), которые соединяются в гонадах (половых железах) и
распределяются по гаметам (сперматозоидам или яйцеклеткам). В каждой гамете,
таким образом, должны присутствовать пангены пальцев, волос, зубов и т. д. Эта
теория, довольно широко распространен-
82
85
семян) с тем или иным признаком. Скрещивая растения по
цвету семян, он получил в поколении F2 6022 желтых семени и 2001 зеленое семя. Скрещивая
растения по окраске цветков, он получил 705 фиолетовых цветков и 224 белых. Эти
цифры еще ничего не говорят, и в похожих случаях предшественники Менделя
опускали руки и утверждали, что ничего разумного по этому поводу сказать
нельзя. Однако Мендель заметил, что отношение этих чисел близко к пропорции
3:1, и это наблюдение подтолкнуло его к простому выводу.
Мендель разработал модель - гипотетическое объяснение того, что
происходит при скрещивании. Ценность модели зависит от того, насколько хорошо
она объясняет факты и предсказывает результаты экспериментов. Согласно модели
Менделя, в растениях имеются некие 'факторы', определяющие передачу
наследственных признаков, причем каждое растение имеет по два фактора для
каждого признака - по одному от каждого родителя. Кроме того, один из этих
факторов может быть доминантным, то
есть сильным и видимым, а другой - рецессивным, или
слабым и невидимым. Желтая окраска семян должна быть доминантной, а зеленая -
рецессивной; фиолетовый цвет доминантен по отношению к белому. Такое свойство
'факторов наследственности' находит отражение в записи генетических
экспериментов: прописная буква означает доминантный признак, а строчная -
рецессивный. Например, желтую окраску можно обозначить как Y, а зеленую как у. Согласно
современной точке зрения, 'факторы наследственности' - это отдельные гены,
определяющие цвет или форму семян, и мы называем различные формы гена аллелями
86
Рис. 4.1. Объяснение результатов,
полученных Менделем.
Каждое растение имеет
две копии гена, определяющего цвет, но передает своим гаметам по одной из этих
копий. Ген Yдоминантен по отношению к гену у,
поэтому семена всех растений поколения F1 с набором генов Yy желтые. В следующем поколении возможны
четыре комбинации генов, три из которых дают желтые семена и одна - зеленые
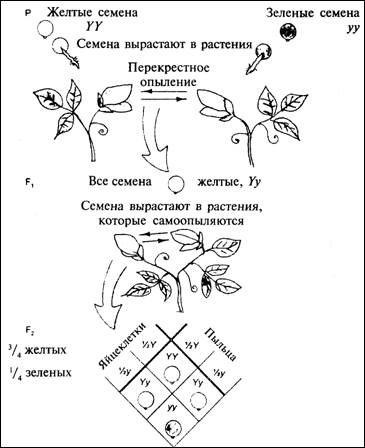
87
94
Заметим, что простая модель, которой мы
руководствовались, основана всего лишь на нескольких довольно разумных
посылках:
♦
каждый организм содержит две копии одного и того же гена;
♦
некоторые аллели могут быть доминантными или рецессивными по отношению к
другим:
♦ при образовании гамет парные гены
расходятся;
♦
при образовании зигот гаметы соединяются случайным образом.
Модель помогает объяснить результаты
экспериментов и исследований, поэтому можно сказать, что она согласуется с
реальностью.
Другой пример: тестеры и нетестеры
Для закрепления пройденного материала
рассмотрим еще один пример. Химики обнаружили вещество фенилтиокарбамид,
которое кому-то кажется горьким, а некоторым - безвкусным. Первых назвали
тестерами, так как они могут обнаружить это вещество. Среди североамериканцев
европейского происхождения около 70% тестеров. Фенотипы тестера и нетестера
определяются наследственностью, причем аллели тестеров доминантны. Обозначим
аллель тестера как Т, а аллель
нетестера как t.
Неудивительно, что у гомозиготных тестеров рождаются тестеры (TT x TT); а у гомозиготных нетестеров - такие же нетестеры (tt х tt). Теперь
представим себе брак между двумя разными гомозиготами:
95
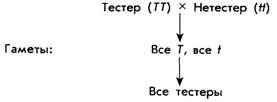
Некоторые скрещивания гетерозигот выявили
следующие результаты:

Самая очевидная комбинация - Tt x Tt. Закон
расщепления Менделя помогает понять, почему получается отношение 3:1. Самый
простой способ наглядно представить, что получается при скрещивании, -
нарисовать так называемую решетку Пеннета, то
есть таблицу, в которой по вертикали и по горизонтали отмечены мужские и
женские гаметы. На пересечении вертикальных столбцов и горизонтальных строк указываются
возможные генотипы.
96
97
ная гомозигота : 2 гетерозиготы : 1
рецессивная гомозигота.
Во всех примерах, которые мы
рассматривали до сих пор, доминантные гомозиготы и гетерозиготы имеют один
общий доминантный фенотип. Однако законы Менделя становятся еще более
наглядными, если гетерозиготы фенотипически отличаются от обоих видов
гомозигот, то есть в случае так называемой неполной
доминантности, при которой фенотип гетерозигот представляет собой
нечто переходное между фенотипами гомозигот. У некоторых видов растений
скрещивание разновидности с красными цветами и разновидности с белыми цветами
дает гибридное потомство с розовыми цветами; далее скрещивающиеся между собой
гетерозиготы дают менделевское отношение 1 красный : 2 розовых : 1 белый. В данном
случае генотипическое отношение 1:2:1 точно соответствует фенотипическому
отношению 1:2:1.
Неплохим уроком по генетике может
оказаться исследование групп крови у людей. Кровь относят к той или иной группе
в зависимости от того, как она взаимодействует с иммунной системой, которая
защищает организм от вторжения таких чужеродных тел, как бактерии и вирусы.
Иммунная система, более всего развитая у птиц и млекопитающих, учитывает
различия в форме молекул, особенно в форме белков. У каждой клетки на
поверхности находится много разных белков и других молекул, причем у каждого
организма свой набор. Он прида-
98
110
Мендель открыл основные законы
наследственности приблизительно в то же время, когда другие биологи принялись
изучать строение клетки. Поэтому когда законы генетики открыли заново в 1900-х
годах, ученые уже смогли донять, что так называемые факторы должны
соответствовать наблюдаемым в реальности структурам - хромосомам. Сейчас мы
знаем, что факторы Менделя - это гены, и они содержатся в хромосомах.
Отождествление абстрактных понятий и реальных структур - крупнейшее достижение
генетики на ранней стадии ее развития, и оно было сделано в основном благодаря
исследованию природы пола.
Клетки и размножение
После того как клеточная теория Шлейдена
и Шванна стала общепринятой, патолог Рудольф Вирхов сделал свой немаловажный
вклад. Он предположил, что не только все организмы состоят из клеток, но и
всякая клетка происходит от клетки-прародителя. Мы уже сказали, что всякая
клетка растет, получая питательные вещества, пока не достигает размера, вдвое
больше обычного, после чего
115
116
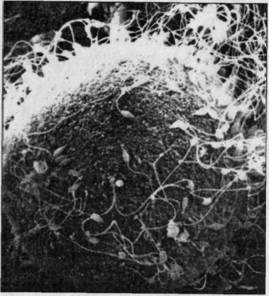
Рис. 5.1. Микрофотография
яйцеклетки, окруженной многочисленными сперматозоидами в момент оплодотворения
установил, что организм развивается в
процессе эпигенеза. Это значит, что части организма формируются постепенно.
Развитие начинается с оплодотворения сперматозоидом яйцеклетки и образования
зиготы. Органы и части тела образуются из соматических клеток (таких, как
клетки мышц, костей и печени); гаметы же - из особых клеток, которые находятся
только в половых железах, семенниках и яичниках. Таким образом, один организм
порождает другой благодаря слиянию и делению клеток, и непрерывную цепь жизни
можно проследить от одного поколения к другому, вплоть до самых далеких наших
предков.
117
Митоз и клеточный цикл
Отдельная клетка растет и делится на две
новые клетки, проходя через клеточный цикл. Цель такого цикла - произвести две
идентичные клетки и более, которые продолжат процесс, получив от родительской
клетки копию ее генома. Так как геном, или совокупность генов, содержится в
основном в хромосомах ядра, клетка для начала должна произвести копии своих
хромосом. Количество хромосом увеличивается вдвое во время С-фазы цикла (от
слова 'синтез'), то есть в тот период, когда реплицируется (удваивается) ДНК и
удваивается геном. Затем, во время митоза (М), хромосомы расходятся, и ядерное
деление дает два одинаковых ядра с идентичным набором хромосом:
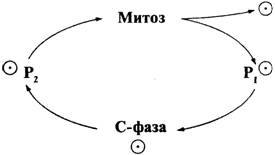
Таким образом, в жизненном цикле клетки
можно выделить два основных события: удвоение хромосом и деление на две
дочерние клетки, каждая из которых получает по одной копии хромосомы.
Между фазами М и С наблюдается
относительно долгая стадия роста (Р1). Между фазами С и М
проходит вторая стадия роста (Р2), когда клетка под-
118
готавливается к митозу. Период,
охватывающий стадии М, P1 и Р2 иногда называют
интерфазой. Этот термин сохранился с того времени, когда ученые имели
представление только о митозе и предполагали, что это самая важная фаза
клеточного цикла. Митоз сам подразделяется на несколько стадий, последовательно
сменяющих друг друга. Более детально процесс репликации, или удвоения ДНК, мы
объясним в гл. 7. Сейчас же отметим, что после прохождения С-фазы все хромосомы
в клетке удваиваются и готовы перейти в дочерние клетки.
Во время интерфазы в ядре мало что
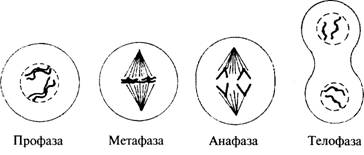
происходит. Когда клетка вступает в первую стадию митоза - профазу (рис. 5.2),
ядерная мембрана распадается, и можно увидеть, что ядро содержит отчетливые
нитевидные хромосомы. На этой стадии каждая хромосома уже удвоена, и обе они
соединены попар-
Рис. 5.2. Стадии митоза:
профаза-хромосомы становятся видимыми, а оболочка ядра распадается
Рис. 5.2. Стадии митоза:
профаза-хромосомы становятся видимыми, а оболочка ядра распадается; метафаза - хромосомы
выстраиваются посреди клетки; анафаза - две
хроматиды каждой хромосомы расходятся к противоположным полюсам; телофаза -
когда образуются новые ядерные оболочки и
хромосомы снова становятся неразличимыми
119
120
клеток другому их поколению. Благодаря
такому же сложному и в высшей степени организованному механизму митоза
отдельная зигота вырастает во взрослый организм, содержащий триллионы клеток.
Клеточное деление не только обеспечивает рост животных и растений, но и
поддерживает здоровье наших организмов. Ежедневно на протяжении всей нашей
жизни митотическое клеточное деление заменяет изношенные клетки кожи новыми,
затягивает наши раны и производит новые красные кровяные тельца.
Зная механизм митоза, можно лучше
рассмотреть хромосомы, которые свободно движутся во время этого процесса.
Поместим каплю крови в пробирку с питательным раствором, в котором могут
размножаться лейкоциты (белые кровяные клетки). Через несколько дней обработаем
раствор колхицином, разрушающим
аппарат веретена деления и останавливающим деление клеток в метафазе, когда их
хромосомы наиболее различимы. Т. С. Сю (Т. С. Hsu) открыл, что, если клетки поместить в раствор с
концентрацией соли, меньшей, чем необходимо, для клеток, они впитывают воду и
разбухают, после чего их хромосомы разворачиваются и распрямляются. Такие
клетки помещают на столик микроскопа так, чтобы их хромосомы были отчетливо
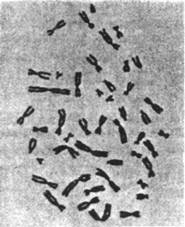
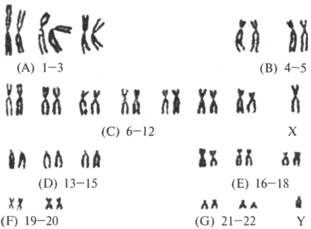
видны, и фотографируют (рис. 5.3а). На фотографии видно, что хромосомы
различаются по длине и форме: одни короткие, другие длинные; их центромеры
занимают различные положения. Каждый био-
121
Рис. 5.3. Для составления
кариотипа делящиеся клетки распределяют на пластине
Рис. 5.3. Для
составления кариотипа делящиеся клетки распределяют на пластине, чтобы их
хромосомы были отчетливо видны, и фотографируют (а). Затем гомологичные
хромосомы на фотографии распределяют по парам и выстраивают по размеру так,
чтобы значительно облегчить их исследование (б)
а
б
122
123
Вспомним, что митоз происходит только при
делении обычных, соматических, клеток и что в результате такого деления в
клетках остается один и тот же набор хромосом. Если бы сперматозоиды и
яйцеклетки образовывались в процессе митоза, то в зиготе было бы в два раза
больше хромосом, чем в клетках родителей, и с каждым поколением количество
хромосом в организме удваивалось бы. Однако этого не происходит. Поскольку у
представителей каждого поколения один и тот же набор хромосом, совершенно
очевидно, что должен существовать еще один вид деления, при котором количество
хромосом уменьшается наполовину. Такое деление называется мейоз. При каждом оплодотворении изначальное
количество хромосом восстанавливается. Половое размножение, таким образом,
можно представить себе как следующий большой цикл событий:

Клетка, у которой только один набор
хромосом, называется гаплоидной (haploos - простой), а клетка с двойным набором хромосом называется диплоидной (diploos - двойной). В кариотипе человека имеется двойной набор 23 хромосом, по-
124
Рис. 5.5. При сперматогенезе из
начальной клетки образуются четыре сперматозоида, а при оогенезе только одна
клетка.
При делении первичного
ооцита один из наборов хромосом переходит в крошечное полярное тельце, которое
может делиться еще раз, но никаких функций не выполняет. При втором делении
образуется еще одно полярное тельце. Так получается большая гаплоидная
яйцеклетка, теряющая незначительную часть начальной цитоплазмы
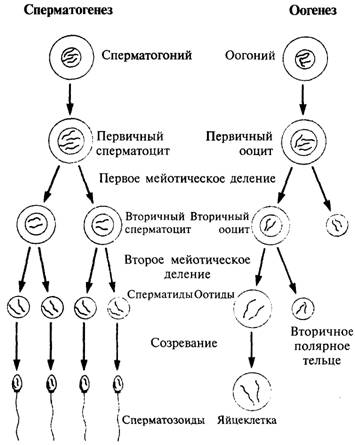
129
запас питательных веществ. В первичном ооците мейоз происходит у границы клетки, и в
конце первого мейотического деления одно дочернее ядро отделяется от основной
клетки в виде полярного тельца. Во
время второго этапа мейоза ядра снова делятся, и одно из ядер ооцита
отделяется. Таким образом, при оогенезе из каждого ооцита образуются три
полярных тельца и одна яйцеклетка.
Взрослый мужской организм ежедневно
производит миллионы сперматозоидов, женский же - одну яйцеклетку за период
приблизительно 28 дней. Поэтому за все время жизни у женщины образуется 400-500
яйцеклеток. Все клетки, способные образовывать яйцеклетки, - около 2 миллионов
- уже содержатся в организме девочки при рождении. К наступлению половой
зрелости количество ооцитов уменьшается до 300 тыс., и лишь малое число
достигает стадии зрелой яйцеклетки.
Мейоз и законы Менделя
В наши дни широко известно, что гены
находятся в хромосомах, хотя в следующем разделе мы постараемся это утверждение
доказать. Рассмотрев процесс мейоза, мы теперь можем найти в нем обоснование
закона Менделя, согласно которому каждая особь содержит два фактора одного
признака, но при этом гаметы переносят лишь один из этих факторов. Каждый
животный организм или растение (у Менделя - горох) содержит диплоидный набор
хромосом, которые переносят два аллеля одного гена. Если вернуться к примеру с
тестерами и нетестерами фенилтио-
130
карбамида, то какая-то из 23 пар наших
хромосом должна переносить либо аллель Т, либо
аллель t. У гетерозигот Tt гомологичные хромосомы в начале мейоза образуют пары,
но во время первой анафазы расходятся, вследствие чего образуются гаметы,
переносящие либо Т, либо t.
Второй закон Менделя, закон независимого
распределения, говорит, что при наследовании двух факторов их распределение
происходит независимо. На самом деле гены распределяются независимо, если они
располагаются на разных хромосомах. Позже (гл. 8) мы столкнемся с так
называемыми сцепленными генами, расположенными на одной хромосоме. В данном
случае важно отметить, что каждая хромосомная пара разделяется независимо от
других. Предположим, что в организме только две пары хромосом: одна переносит
аллели А и
а, а другая - аллели В и b. У двойных гетерозигот АаВb в метафазе I
все хромосомы выстраиваются парами
посередине клетки:
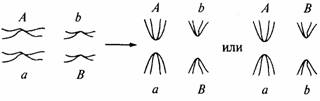
В анафазе I может
получиться так, что аллели А и В попадут в одну дочернюю клетку, а аллели а и b - в другую. Но существует равная вероятность того, что в одной дочерней
клетке окажутся аллели А и b, а в другой - аллели а и В. Многократное мейотическое деление даст
гаметы всех четырех ти-
131
пов: AB, Ab, aB и ab. Это
приводит к той модели наследственности, которую мы изучали в гл. 4.
Основные процессы, происходящие при
мейозе и митозе, были изучены к концу XIX века.
Теперь известно, что это довольно сложный механизм распределения хромосом по
дочерним клеткам, но до начала ХХ века ученые не понимали их биологической
роли. И только после повторного открытия законов Менделя в 1900-х годах Теодор
Бовери и Уолтер Саттон независимо друг от друга провели аналогию между
поведением гипотетических факторов и хромосом при мейозе. В 1902 году они
выдвинули хромосомную теорию наследственности, заявив, что наследственные
факторы Менделя располагаются на хромосомах. Они указали на следующие
параллели:
Однако такие параллели в поведении
хромосом и генов еще не являются доказательством того, что
132
гены расположены в хромосомах, и многие
биологи, в том числе генетики, поначалу отзывались о теории Бовери-Саттона
крайне скептически. Тем не менее различные эксперименты и наблюдения,
проведенные в первые десятилетия ХХ века, подтвердили эту гипотезу.
Еще в древности люди заметили, что
некоторые заболевания появляются почти исключительно у мужчин, хотя передаются
по материнской линии. Самый известный пример - гемофилия,
или недостаточная свертываемость крови. Древние евреи запрещали
проводить обязательное обрезание ребенку, если два его старших брата умирали от
кровотечения, то есть признавали наследственный характер этого заболевания. К XII веку ученые-раввины признали, что гемофилия поражает
мужчин, хотя, что парадоксально, передается по женской линии. Чарльз Дарвин
также обратил внимание на этот особый вид наследования. В 1875 году он описал
историю одной индийской семьи, в которой у 10 мужчин на протяжении четырех
поколений были очень мелкие зубы, слабый волосяной покров, раннее облысение и
ненормально сухая кожа. Ни у одной женщины в семье эти признаки не проявились,
хотя они и передавали все эти особенности своим сыновьям. В то же время ни у
одного из мужчин не родились дети с подобными признаками.
Такой характер наследования объясняется
различием в половых хромосомах, отчетливо наблюдаемым в кариотипе: у женщин две
Х-хромосомы,
133
|
Гены
|
Хромосомы
|
|
1. Содержатся в организме парами, по одному от каждого родителя.
|
1. Содержатся гомологичными парами в диплоидных клетках в результате объединения гамет каждого родителя.
|
|
2. Аллели парных генов расходятся по разным гаметам.
|
2. Во время мейоза расходятся по разным гаметам.
|
|
3. Разные пары распределяются независимо друг от
друга.
|
3. Разные пары распределяются независимо друг от друга.
|
тогда как у мужчин одна такая хромосома и
одна Y-хромосома поменьше, которая ведет себя как
гомологичная Х-хромосоме при мейозе. Во всех яйцеклетках содержится только
Х-хромосома, тогда как в половине сперматозоидов находится Х-хромосома, а в
другой половине - Y-хромосома. Это объясняет, почему
мальчиков и девочек рождается примерно поровну: ведь если яйцеклетку
оплодотворит сперматозоид с Х-хромосомой, то получится женская зигота ХХ, а
если сперматозоид с Y-xpoмосомой, то мужская зигота XY. Забавно, что пол ребенка определяют сперматозоиды,
хотя до сих пор именно женщин часто винят в том, что они родили ребенка не того
пола. Бывало так, что цари и короли разводились с женами, которым не удавалось
родить им наследника. Однако у некоторых других животных, включая земноводных,
птиц и бабочек, пол определяет яйцеклетка; у мужских особей этих животных две
идентичные Z-хромосомы, а у женских особей -
две разные: W- и Z-хромосомы.
В Y-хромосоме человека содержится мало известных генов.
Небольшой участок этой хромосомы под названием SRY определяет
развитие семенников (яичек) вместо яичников; любая зигота с Y-xpoмосомой
развивается в мужской организм, а зигота без нее - в женский. Любой признак,
определяемый геном на Y-хромосоме,
должен передаваться от отца к сыну и проявляться в любом случае. Почти
единственный подтвержденный признак такого рода - так называемые волосатые края
уха. Правда, этот признак проявляется в жизни поздно и по-разному, поэтому
механизм его наследственности
134
выяснен не совсем точно, хотя похоже, что
он должен передаваться по мужской линии.
Наряду с этим многие признаки передаются
с Х-хромосомой, и их местонахождение определяется благодаря особому образцу
наследования. Хороший пример - дальтонизм по красному и зеленому цветам.
Обозначим Х-хромосому с мутантным аллелем как XС,
а хромосому с нормальным аллелем как Х+. Поскольку мутация
дальтонизма рецессивная, то у женщины, гетерозиготной по этому гену (ХСХ+),
зрение нормальное. Но у мужчины, получившего мутантный ген с единственной
Х-хромосомой (XСY), нет нормального аллеля, потому проявляется
рецессивный аллель. Мужчина-дальтоник передает свою ХС-хромосому
всем своим дочерям, и они (как правило) становятся гетерозиготными
носительницами этого признака. Женщина с равной вероятностью может передать
своим сыновьям как XС-, так и Х+-хромосому.
Женщины-дальтоники встречаются редко, потому что они должны унаследовать обе
ненормальные хромосомы от дальтоника-отца и от гетерозиготной матери;
вероятность такого сочетания будет равна 50%.
Признаки, сцепленные с Х-хромосомой, в
родословных распознавать легко, потому что женщины передают их приблизительно
половине сыновей, а от отцов они передаются через дочерей внукам и проявляются
через поколение. Такой характер наследования присущ нескольким сотням признакам,
включая некоторые виды облысения и мышечную дистрофию Дюшенна. Один из самых
известных случаев - передача по наследству гемофилии в родословной европейских
монархов (гл. 14).
135
Обычно мужчины и женщины имеют хорошо
выраженный фенотип, определяемый их набором хромосом - XY или ХХ. Но
иногда рождаются дети с необычным числом половых хромосом, и это происходит в
результате ненормального развития гамет. Два подобных синдрома названы по
именам первых описавших их врачей. Синдром Клайнфельтера проявляется у
мальчиков, которые обычно высокие, с гинекомастией (развитие молочных желез по
женскому типу), пониженным умственным развитием и маленькими яичками. В 1959
году Якобс и Стронг установили, что синдром Клайнфельтера связан с наличием
лишней Х-хромосомы, то есть с набором хромосом XXY.
Другой случай ненормального развития
гамет называется синдромом Тернера и проявляется у девочек. У них нет яичников,
они невысокие, с недоразвитыми вторичными половыми признаками (маленькая грудь).
Хромосомный набор у таких девочек - Х0, то есть одна Х-хромосома (0 обозначает
отсутствие хромосомы). Поскольку такие женщины гомозиготны по Х-хромосоме, у
них проявляется рецессивный фенотип, например дальтонизм, обычно свойственный
мужчинам. Рождение одного ребенка с генотипом XXY приходится
приблизительно на каждые 700 рождений, а с генотипом Х0 - на каждые 2500. Кроме
того, на каждую 1000 рождений приходится один случай XXX; эти девочки внешне нормальные, хотя и с некоторыми
недостатками умственного развития.
Как же возникают такие случаи?
Непосредственная причина пока еще не известна, но ясно, что во
136
Рис. 5.6. Последствия
нерасхождения Х-хромосом в ооците на первом этапе мейоза и оплодотворения
сперматозоидами с хромосомами X или
Y.
Нерасхождение на втором
этапе мейоза (не показано)
может привести к еще большему увеличению числа Х-хромосом
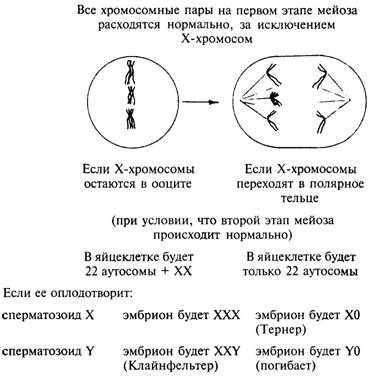
время мейоза эти хромосомы не расходятся
как следует (рис. 5.6). Такое явление и называется нерасхождением
хромосом. Оно происходит в гаметах каждого пола, на первом или
втором этапе мейоза или сразу на двух этапах. В результате нерасхождения
образуются гаметы с двумя половыми хромосомами (ХХ, YY, XY) или вовсе
без половой хромосо-
137
142
хромосомным набором и фенотипом. Если
лишние или недостающие хромосомы влияют на выраженность гена, то гены и в самом
деле расположены в хромосомах.
Попытки определения пола
В древности интерес к вопросам размножения и воспроизводства подогревался желанием предсказать пол детей. На протяжении многих столетий люди предпочитали, чтобы у них рождались мальчики. Так как во многих культурах считалось, что наследственные признаки передаются исключительно через мужское семя, то полноправными наследниками отца признавали только сыновей. Если человек надеялся на память потомков, если он желал оставить после себя воспоминания о своей жизни, то у него обязательно должны были рождаться мальчики. Во многих цивилизациях собственность передавалась по наследству только мужчинам, и, естественно, мужчинам хотелось передать все свое накопленное имущество детям, а не каким-то дальним родственникам. Там, где исключительное положение в обществе занимали цари, короли, вожди, шаманы или жрецы, все члены общества также надеялись на то, что у этих высокопоставленных лиц родятся наследники, и тем самым удастся избежать беспорядков. Иногда люди надеялись на то, что их сыновья удачно выберут себе жену, и та принесет в их дом богатое приданое. Другие мечтали о сильных воинах и искусных охотниках, которые прославили бы их род и обеспечили процветание всем членам семьи. Дочери же часто оказывались 'лишними ртами', и, кроме того, во многих цивилизациях требовалось давать им приданое, что могло разо-
Продолжение на след. странице
143
рить семейство, где рождалось несколько дочерей подряд. По этим и многим другим причинам рождение сына воспринималось как нечто очень важное и значительное. Подобное отношение к мальчикам сохраняется и поныне.
Вместе с тем в истории человечества были и такие общества, где особое внимание уделялось дочерям, хотя порой их считали всего лишь ценной собственностью, которая передается от одного наследника другому. В других обществах наследование шло по женской линии, и тогда рождение дочери воспринималось как истинный праздник. Но вне зависимости от того, по какой линии шло наследование в том или ином обществе, во всех обществах и цивилизациях бытовали свои, часто очень любопытные, 'правила' определения пола будущего ребенка, своего рода 'первобытная генная инженерия'.
В Древнем Египте не было указаний, как зачать ребенка нужного пола, но правила определения пола имелись. Египтяне считали, что если мочой беременной женщины каждый день поливать семена пшеницы и ячменя, то пшеница прорастет, если у нее будет мальчик, а ячмень прорастет, если будет девочка. Если же ничего не прорастало, это означало, что женщина не беременна. У этого странного на первый взгляд метода есть некоторые основания, потому что моча беременных женщин содержит химические вещества, которые действительно способствуют прорастанию определенных растений.
Древние евреи желали, чтобы у них рождались мальчики, так как в их обществе женщины обладали низким статусом и не наследовали имущество. Они, например, считали, что если мужчина сожжет зерно соседа, то у него не будет наследников, но если, как сказано в Талмуде, он поставит свое ложе с севера на юг, то у него, скорее всего, родятся мальчики.
144
Евреи разделяли с арабами и индусами веру в то, что рождение мальчика - настолько счастливое и радостное событие, что если беременную женщину не тревожить и не огорчать, то у нее обязательно родится мальчик.
Считалось также, что пол ребенка зависит от питания матери. В древнеиндийской 'Атхарваведе' (написанной до 700 г. до н. э.) содержится предписание пить на четвертый день менструации 'гарбхакарану', то есть напиток из вареных семян риса и кунжута. В таком случае якобы обязательно родится мальчик. В древних текстах также можно встретить рецепты получения нужного оттенка кожи у сыновей, а также рецепты, которые должны ему помочь в будущем владеть тем или иным количеством 'Вед'.
Во времена Гомера греки радовались рождению дочерей. Сыновья часто подвергались опасностям и погибали, тогда как дочерям доставались скот и другое имущество. Но несколько столетий спустя наследовать собственность могли уже только сыновья. Дочерям давали только приданое, когда они выходили замуж. Если мужчина умирал, не оставив после себя наследников, его собственность вместе с женой переходила к ближайшему родственнику.
Естественно, что люди пытались заранее определить пол своего ребенка. Гиппократ учил, что у женщины со здоровым цветом кожи родится мальчик. Он также полагал, что если зародыш двигается уже на третьем месяце беременности, то такой энергичный ребенок обязательно должен быть мальчиком. Если лицо женщины покроется веснушками, то она родит девочку (у других народов веснушки предвещают рождение мальчика).
Гиппон Регийский полагал, что пол ребенка определяет отец: густое семя отца означает, что родится
Продолжение на след.
странице
145
мальчик, а жидкое - девочка. Для того чтобы обеспечить рождение мальчика, греки вступали в половое сношение при северном ветре, потому что северный ветер, 'как известно', уплотняет и усиливает семя. Для девочек же подходящим оказывался южный ветер, который разбавляет и ослабляет семя. Эмпедокл, напротив, считал, что рождение мальчиков зависит от женщин, у которых должна быть особо теплая матка.
Многие теории пола коренятся в древнем убеждении, что правая сторона тела более важна, чем левая, а такое убеждение в свою очередь объясняется тем, что большинство людей правши. Часто считали, что пол ребенка определяет семя из одного или другого мужского яичка. Иногда мужчины, для того чтобы зачать мальчика, перевязывали одно из своих яичек. Аристотель возражал против такого обычая, но он сохранялся долго; даже в XVIII веке некоторые французские аристократы удаляли левое яичко в надежде зачать наследника.
Большое значение придавалось и снам женщины. В Индии считалось, что если женщине приснится мужская еда, то у нее будет мальчик. В некоторых славянских странах то же самое означают ножи и дубинки, а сны об источниках и гулянках означают рождение девочки. В некоторых областях Германии мужчины клали в кровать топор, желая зачать ребенка мужского пола, и оставляли его в сарае, когда хотели зачать девочку. В Японии X века женщины, надеясь 'взбодрить' ребенка и сделать из него мальчика, отправлялись на охоту или сражение. Среди японцев также бытовало забавное убеждение, что если мужчина позовет свою жену, когда та справляет естественные потребности и при этом повернется налево, то у нее родится девочка. Древнее китайское правило гласит: если на седьмом месяце беременно-
сти мать почувствует, как ее ребенок двигает рукой влево, то это будет мальчик; если же ребенок на восьмом месяце двигает рукой вправо, то это будет девочка.
Многие люди и поныне разделяют различные предрассудки относительно пола ребенка и верят в то, что пол можно определить по частоте сердцебиения, по тому, насколько часто будущая мать испытывает тошноту, по качеству ее молока. В наши дни пол ребенка возможно совершенно точно определить медицинскими методами, а в будущем, вероятно, станет возможным и выбирать пол ребенка при зачатии. Раньше люди не сомневались, что возможность определить пол ребенка - это обязательно благо, но теперь, когда на нас неожиданно легла такая большая ответственность, мы не всегда уверены в том, что сможем с ней справиться.
Глава 6. ФУНКЦИЯ ГЕНОВ
Итак, мы уже рассмотрели, как хромосомы и
гены передаются от поколения к поколению, но еще не узнали, за что именно они
отвечают и как функционируют. Пора подробнее осветить этот вопрос.
Гены и нарушения метаболизма
Люди - плохой 'материал' для изучения
законов наследственности, потому что у них трудно получить достаточно надежные
данные, но первые наблюдения, как гены осуществляют свою функцию, были сделаны
именно на людях. В самом начале ХХ века английский врач Арчибальд Гаррод1 заинтересовался
наследственностью лиц с определенными нарушениями и начал сравнивать их
биохимические анализы с анализами нормальных людей. Одно из таких нарушений,
передающееся по аутосомному рецессивному образцу, - алкаптонурия,
выражающаяся, в частности, в том, что моча таких людей на воздухе
темнеет (становится темно-красной или черной). Обычно это замечают уже
родители, когда видят, что пеленки их ребенка становятся темными. Люди с этим
нарушением выде-148
ляют большое количество гомогентизиновой
кислоты (алкантона), которая и темнеет при реакции с кислородом. У нормальных
людей клетки преобразуют эту гомогентизиновую кислоту в малеилуксусную кислоту.
Отсюда следует, что алкаптонурия происходит от отсутствия ферментов, которые
обычно ускоряют эту реакцию. Вспомним, что все вещества в организме образуются
или разрушаются в процессе метаболических реакций, или метаболических путей,
которые представляют собой серию химических реакций при участии катализирующих
ферментов. Гаррод назвал алкаптонурию и схожие с ней симптомы врожденными
нарушениями метаболизма.
Характер этих нарушений предполагает, что
гены передают информацию о производстве ферментов. У обычных людей имеется
аллель Al, который вырабатывает
фермент, преобразующий гомогентизиновую кислоту в малеилуксусную кислоту.
Страдающие алкаптонурией гомозиготны по рецессивному аллелю al, содержащему 'испорченную' информацию. У
гетерозигот также содержится один нормальный аллель, поэтому у них не
проявляется признаков нарушения метаболизма. Гаррод заметил также, что
альбинизм проявляется похожим образом: в коже, волосах и радужной оболочке глаз
отсутствует обычный пигмент меланин. Меланин образуется с помощью ферментов из
тирозина в ходе нескольких реакций, у альбиносов не вырабатывается фермент для
одной из них. Такая связь между генами и составляющими веществами организма
весьма знаменательна.
Нам известно несколько наследственных
нарушений, блокирующих метаболические пути, веду-
149
щие к образованию меланина и
малеилуксусной кислоты (рис. 6.1). На основании подобных наблюдений можно
сделать вывод, что гены ответственны за выработку ферментов и как следствие
контролируют метаболизм в целом. Некоторые нарушения метаболизма даже
определили ход истории. Например, известно такое нарушение метаболизма, как
порфириновая болезнь, при которой не расщепляются порфирины. Их избыток
приводит к пси-
Рис. 6.1. Метаболические пути
тирозина и фенилаланина в организме человека.
Отсутствие ферментов на
определенных этапах приводит к специфическим наследственным нарушениям
метаболизма
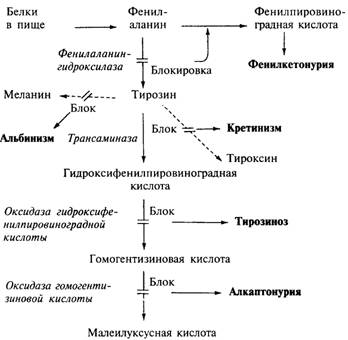
150
хическим расстройствам. Несдержанное
поведение английского короля Георга III -
один из факторов, ускоривших революцию в Америке - было результатом
порфириновой болезни, унаследованной им от шотландской королевы Марии.
Гены и ферменты
В 1944 году Джордж Бидл и Эдвард Тэйтем
подтвердили правильность выводов Гаррода на примере хлебной плесени Neurospora (эта плесень ярко-оранжевого цвета иногда образуется
на черством хлебе). Хлебную плесень можно выращивать на простой питательной среде - в смеси сахара, витамина
биотина и нескольких солей, растворенных в воде. Из этих веществ плесень
синтезирует все необходимые сложные компоненты. Такие организмы, не требующие
для своей жизнедеятельности сложных веществ, называются прототрофами. Бидл и Тэйтем подвергали плесень
облучению с целью ускорения процесса мутаций и затем искали среди различных
популяций ауксотрофы, то есть
организмы, утратившие способность синтезировать одно из веществ, необходимых
для их роста, и, таким образом, способных жить только в питательной среде с
этим веществом.
Особое внимание Бидл и Тэйтем обратили на
мутантную плесень, которая для своего существования требовала аминокислоту
аргинин. Аргинин - необходимая составляющая всех белков, и обычная плесень Neurospora синтезирует ее из сахара и других веществ раствора.
Однако нарушения метаболического процесса у мутантной плесени не позволя-
151
167
рекции многих наследственных заболеваний
станут эвфенические меры.
Успех эвфеники зависит от изменения
окружающей среды. И тут на первый план выходит мнение, что большинство
заболеваний вызывается внешними, а вовсе не наследственными факторами. Кроме
того, степень проявления признаков, характерных для многих наследственных
нарушений, связанных с одним геном (и для некоторых, связанных с многими генами),
во многом зависит от внешних условий. Поэтому, как выразилась медицинский
генетик Патрисия Бэйрд, не следует переоценивать генетический подход применительно
к общественному здравоохранению. (Как мы увидим в гл. 12, такой подход будет
приобретать все большую значимость по мере прогресса в геномике.) Пока
наибольших достижений в области здравоохранения можно ожидать на уровне
улучшения и контроля внешней среды.
Глава 7. НАСЛЕДСТВЕННЫЙ МАТЕРИАЛ, ДНК
С самых первых научных исследований
механизма наследственности ученых не переставал интересовать главный вопрос:
'Что представляет собой наследственный материал?' В начале ХХ века гипотеза
Саттона-Бовери о том, что гены находятся в хромосомах, стала общепризнанным
мнением. Но какое химическое вещество в хромосомах служит переносчиком
генетической информации? Еще на заре биохимии ученые предполагали, что на роль
носителя информации подходят два основных вида химических веществ клетки -
белки и нуклеиновые кислоты. И хотя об их строении было известно мало, белки,
как более сложные, казались наиболее подходящим кандидатом. Поэтому считалось,
что гены состоят из белков. Вместе с тем некоторые опыты свидетельствовали о
том, что не стоит сбрасывать со счетов и нуклеиновые кислоты. Когда Э. Б.
Уилсон публиковал свой классический труд 'Клетка и ее роль в наследственности и
эволюции', в одном издании он написал, что наиболее важный материал - белки, а
в другом издании назвал нуклеиновые кислоты. Однако никто ничего не знал
наверняка.
Ответ на этот ключевой вопрос дали
исследования бактерий и поражающих их вирусов. В неболь-
169
177
особо не вспоминали, в основном благодаря тому, что антибиотики, широко распространившиеся во время Второй мировой войны, позволяли легко излечивать инфекционные заболевания. Однако в Восточной Европе, особенно в Польше и Советском Союзе, фаговой терапии продолжали уделять важное внимание. В течение последних нескольких лет о ней вновь заговорили, потому что многие патогенные бактерии выработали иммунитет ко всем применявшимся против них антибиотикам. Так фаговая терапия снова входит в моду. Но это совсем другая история, и она еще ждет своего продолжения.
В 1915 году англичанин Фредерик Творт и
канадец Феликс Д'Эрелль независимо друг от друга открыли бактериофаги, которые вызывают инфекции среди
бактерий. Сама идея об инфекциях среди бактерий может показаться на первый
взгляд странной, поскольку мы уверены, что инфекции вызывают сами бактерии.
Однако биологам хорошо известно, что у многих паразитирующих организмов имеются
свои, более мелкие паразиты. Бактериофаги, или, как еще их называют, фаги, представляют собой вирусы, растущие
внутри бактерий, подобно тому как другие вирусы размножаются внутри клеток
растений или животных. На их примере можно исследовать общие особенности
поведения вирусов.
Прежде всего, вирусы - это не организмы.
Каждый организм состоит из одной или нескольких клеток, но полностью
сформированные вирусные частицы, вирионы, гораздо
меньше любой клетки. Они
178
состоят из нуклеиновой кислоты (ДНК или
РНК), заключенной в защитную белковую оболочку (рис. 7.3). Большинство вирионов
имеют сферическую или вытянутую форму. Вирусы животных обычно попадают внутрь
клеток, где присоединяются к поверхности клетки и поглощаются (подобно
питательным веществам); вирусы растений обычно проникают через порезы,
сделанные насекомыми или червями. Многие фаги, как тот, что показан на рис.
7.3, прикрепляются к бактериям при помощи 'хвоста', или отростка. Это можно
было увидеть еще на первых электронных микрофотографиях, сделанных до 1945
года. Тогда же выяснили, что фаги приблизительно наполовину состоят из белка и
наполовину из ДНК. И хотя больше почти ничего о вирусах не знали, они оказались
прекрасным материалом для генетических исследований.
Рис. 7.3. Общее строение вирусных
частиц, или вирионов, вирусов бактерий и вирусов животных

179
Рис. 7.4. Фаги выращивают в чашках
Петри, смешивая их образец с бактериями, в которых они могут размножаться, и с
агаром.
Эту смесь наливают
поверх питательного агара в чашку. За несколько часов бактерии размножаются и
равномерно распределяются по всему верхнему слою. Чистые участки, или бляшки,
указывают на те места, где началась инфекция фагов, убивших бактерии

С фагами удобно работать еще и потому,
что они очень быстро размножаются, и за полчаса образуется 100-200 новых фагов.
Их также легко выращивать в чашках Петри (рис. 7.4); немного фагов смешивают с
теплым агаром и бактериями, и эту смесь наливают поверх слоя питательного
агара. Бактерии растут тонким равномерным слоем, который называется газоном, а деятельность фагов приводит к
образованию стерильных участков, или бляшек, в
центре которых начиналась инфекция, погубившая клетки вокруг. Определить
количество фагов в любом материале можно по количеству бляшек в образце. Кроме
того, размер и форма бляшек обычно указывают на те или иные особенности фага.
180
182
Рис. 7.5. Схема протекания фаговой
инфекции
Рис. 7.5. Схема
протекания фаговой инфекции: Л - инфицирующий фаг внедряет свою ДНК в клетку, и
эта ДНК начинает преображать клетку в фабрику по производству очередных фагов;
ферменты, производимые фаговой ДНК, разрушают ДНК клетки-хозяина и
останавливают действие ее генов; В - фаговая ДНК многократно реплицируется;
происходит синтез новых фаговых капсидов; С - новые белки капсидов объединяются
с ДНК и образуют новые фаговые частицы; D - новые фаговые частицы выходят после
разрушения клетки
ко минут фаговая ДНК начинает
синтезировать новые фаговые белки. Вначале создаются белки, отключающие функции
клетки-хозяина; некоторые из них прекращают синтез белков хозяина, а другие
являются ферментами, расщепляющими ДНК хозяина. Ряд других ферментов начинает
репликацию ДНК фага. Вскоре синтез этих ферментов прекращается. Затем
включаются новые гены, которые руководят строительством белков для капсида - белковой оболочки вируса. Эти белки
самопроизволь-
183
но собираются в новые куски капсидов -
головки, отростки и нити. Другие ферменты упаковывают вирусную ДНК в головку.
Примерно через 30 минут после заражения клетку обычно заполняет две сотни новых
вирионов, после чего происходит ее разрушение, или лизис,
под действием других ферментов фага.
Вспомним, что основными строительными
компонентами организма служат полимеры. Нуклеиновые кислоты - это тоже
полимеры, хотя они сильно отличаются по своему строению от белков. Их еще
называют полинуклеотидами, потому что
они состоят из мономеров - нуклеотидов. Нуклеотид
состоит из трех частей: основания, связанного
с сахаром, который, в свою очередь,
связан с фосфатом (РO4).
Нуклеиновая кислота называется по сахару, который входит в ее состав;
рибонуклеиновая кислота (РНК) содержит рибозу, а дезоксирибонуклеиновая кислота
(ДНК) содержит дезоксирибозу (в которой кислорода на один атом меньше).
Основания представляют собой большие кольцевые молекулы с атомами азота;
нуклеотиды ДНК имеют одно из четырех оснований: аденин,
гуанин, цитозин и тимин (обозначаются
A, G, С и Т; в РНК
тимин заменяет урацил - U). Цитозин, тимин и урацил имеют по одному кольцу
атомов и называются пиримидиновыми основаниями; аденин
и гуанин имеют по два кольца и называются пуриновыми
основаниями. Атомы углерода и азота в кольцах для удобства
обозначают порядковыми номерами; атомы углерода сахара - от 1' до 5'.
184
186
кой книге 'Двойная спираль'. Чтобы
получить более объективную информацию о ходе работ, эту книгу, пожалуй, лучше
читать вместе с книгой Анны Сейр 'Розалинда Франклин и ДНК'.
Основная догадка Уотсона и Крика
заключалась в том, что главная роль в структуре ДНК принадлежит основаниям, и
надо как-то учитывать правило Чаргаффа: А = Т, G = С. Они
предположили, что молекула ДНК состоит из двух полинуклеотидных цепей с
противоположной полярностью, закрученных друг относительно друга по спирали (рис.
7.6). Между собой эти цепи удерживаются посредством оснований, соединенных
попарно, при-
Рис. 7.6. Форма двойной спирали
ДНК, по Уотсону и Крику
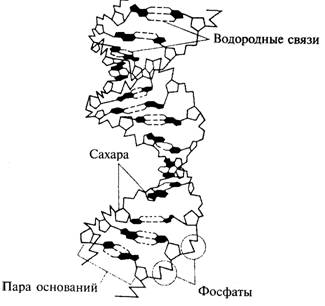
187
чем аденин может соединяться только с
тимином, а гуанин - только с цитозином:
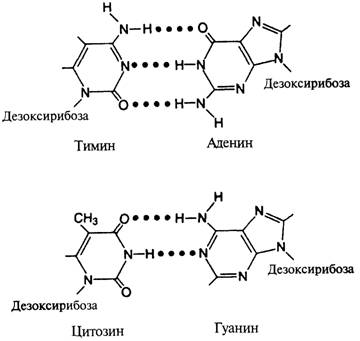
Между основаниями существуют слабые водородные связи, в которых слегка
отрицательно заряженные атомы О и N связаны между собой посредством водорода (имеющим
небольшой положительный заряд). Основания, которые связываются друг с другом,
называются комплементарными друг
другу; это значит, что их форма соответствует друг другу, как рука
соответствует перчатке или ключ - замку. Именно комплементарность оснований
определяет механизм наследственности, а через него и все основные законы
биологии. Модель Уотсона и
188
189
Рис. 7.7. При репликации ДНК
комплекс ферментов разъединяет цепи двойной молекулы
Рис. 7.7. При репликации
ДНК комплекс ферментов разъединяет цепи двойной молекулы, и каждое открытое
основание привлекает к себе комплементарный нуклеотид. Этот процесс
продолжается, пока из двух цепей не вырастут две идентичные молекулы
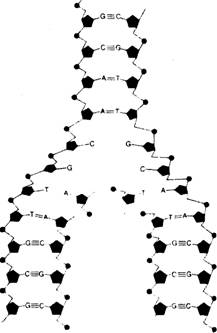
процесса. В действительности все
происходит гораздо сложнее, особенно если учитывать, что цепи ДНК могут расти
только с 3'-конца. Суть процесса сводится к тому, что молекулы ДНК-полимеразы
движутся вдоль каждой цепи и синтезируют комплементарные цепи, образуя таким
образом двойную спираль вместо одинарной. Каждое свободное основание
связывается исключительно с компле-
190
Глава 8. СТРОЕНИЕ ГЕНА
Итак, мы уже знаем, что ген - это
фрагмент ДНК, расположенный на хромосоме и определяющий структуру полипептидной
цепи. В гене могут происходить мутации, и о том, что гены существуют, мы узнали
во многом именно благодаря мутациям. Мутации часто приводят к потере фермента,
поэтому мы знаем, что существуют нормальные гены и нормальные ферменты, а также
то, что гены передают информацию о постройке ферментов следующему поколению
организмов. Остается только выяснить, как устроены сами гены и как они
функционируют. В этой главе мы покажем, как с помощью генетического анализа
можно подробно изучить строение генов и понять, как с их помощью в клетке
создаются белки.
То, что гены расположены в хромосомах,
казалось бы, не соответствует тому факту, что у людей только 23 пары хромосом и
вместе с тем тысячи различных признаков, которым должны соответствовать тысячи
различных генов. Одних только при-
196
знаков, сцепленных с Х-хромосомой,
несколько сотен, а на самой короткой аутосоме расположены также сотни генов.
Как это согласуется с менделевским законом независимого распределения
признаков? Это значит, что закон независимого распределения признаков применим
только для генов, расположенных на разных хромосомах; сначала ученым необходимо
было определить основные законы и выяснить природу наследственности на примере
простейших признаков. На самом деле многие гены расположены на одной и той же
хромосоме, поэтому они, как правило, наследуются вместе. Такие гены называются сцепленными. Одно из достижений современной
генетики и заключается в том, что созданы карты
сцепления для многих признаков. На этих картах показано также
относительное положение генов на хромосомах, и мы увидим, что эти карты имеют
не только теоретическое, но и практическое значение.
Место, которое ген занимает на хромосоме,
называется локусом. За исключением
тех редких случаев, когда происходит перестройка хромосомы, у всех представителей
отдельного биологического вида каждый ген имеет строго определенный локус. Мы
уже говорили, что о существовании генов узнали по мутациям, которые обычно
изменяют гены, делая их дефектными или необычными. Большинство наследственных
признаков известны по таким наследственным заболеваниям, как гемофилия,
дальтонизм и фенилкетонурия. Нормальные аллели гена называются дикими, хотя, как правило, этот термин
применим только для некоторых организмов, с которыми проводят опыты. Гены,
определяющие такие признаки человека, как цвет глаз или группу
197
202
Рис. 8.1. Генетическая карта
плодовой мушки Drosophila melanogaster
нотип A Ht/a ht, а у нормальной матери генотип ht a/ht а. На основании этих данных можно уточнить вероятность передачи дефектного
гена. Ребенок, гомозиготный по а, с вероятностью в 95% не будет иметь Ht, потому что Ht у
больного родителя связан с А и может
отделиться лишь в 5% случаев. Точно так же и для ребенка с аллелем А существует
203
204
До середины 1940-х годов ученые полагали,
что гены, скорее всего, представляют собой хромомеры, то есть крохотные комочки
вдоль хромосом, благодаря которым хромосомы похожи на цепочки бус, и что
кроссинговер происходит только между генами.
Но некоторые опыты на плодовой мушке Drosophila melanogaster показали, что кроссинговер может происходить и внутри
гена. Предположим, что в каком-то локусе двух гомологичных хромосом
располагаются два явно выраженных мутантных аллеля; у мушки могут быть разные
аллели, так что мушка гетерозиготна пр этим аллелям. У таких мух мутантный
фенотип, потому что обе копии гена мутировали. Но иногда такие мушки дают
нормальное, 'дикое', потомство, которое могло бы появиться только в результате
рекомбинации. Это значит, что ген представляет собой не неделимый кусок
хромосомы, а линейную последовательность вдоль хромосомы и что различные аллели
гена могут возникать в результате мутаций во многих местах этой
последовательности, а между различными участками гена возможны рекомбинации.
Обозначим два аллеля цифрами 1 и 2, а
их нормальные ('дикие') участки - знаком плюс. Для наглядности каждый 'дикий'
участок расположим напротив мутантного. Гетерозиготные по обоим аллелям мушки
имеют следующий генотип:
с промежутком между двумя участками
одного гена, где может происходить кроссинговер (очень
205
Макс Дельбрюк выбрал для своих
исследований фаги, потому что они представляют собой очень простую
биологическую систему: крохотные частички, которые могут воспроизводить себе
подобных в других клетках и, как предполагалось, переносить
206
некий генетический материал. Первый
серьезный эксперимент с фагами провел Херши, доказав, что различные штаммы фага
Т2 могут рекомбинировать. Для этого ему, конечно, необходимо было выделить
генетически разные штаммы, и первые обнаруженные им мутанты отличались формой
стерильных пятен. Например, один из мутантов образует крупные пятна с четкими
краями, и Херши обозначил его буквой r (от англ. rapid - быстрый, то есть быстро лизирующий
мутант); мутанты tu (turbid - мутный) образуют мутные пятна; а мутанты mi (minute - мелкий) - очень маленькие пятна. Все эти мутанты имеют отчетливо
выраженный фенотип, то есть легко обнаружить образованные ими пятна, выделить
их и вырастить штамм фагов с генотипом, отличающимся от дикого.
Бактерии можно заразить несколькими
фагами одновременно. Херши заражал клетки мутантами r и tu, взятыми
в достаточном количестве, чтобы почти каждая клетка была заражена фагами обоих
типов. Большая часть потомства этих фагов принадлежала к типам r или tu, но
появлялось также некоторое количество двойных мутантов r, tu и диких фагов. Таким образом, взаимодействовать могут
даже ДНК вирусов, образуя в процессе кроссинговера рекомбинации. Херши
использовал в своих экспериментах несколько независимых мутантов и, приняв
частоту рекомбинаций между ними за условное относительное расстояние (как в
классической генетике), смог расположить участки их мутаций на генетической
карте. С тех пор эта карта была дополнена и расширена.
207
Тонкая структура гена
Сеймур Бензер исследовал тонкую структуру
гена с помощью фагов Т4, среди которых ему удалось выделить редкие внутригенные
рекомбинанты. Бензер сосредоточил внимание на классе мутантов r - rII. Они растут и образуют большие стерильные пятна на штамме Е. coli В, но не растут на штамме Е. coli К. В отличие от них дикие формы rII+ растут
и на В, и на К. Бензер обнаружил сотни новых мутантов rII, которые оказались полезными не только для составления карты, но и для
уточнения того, что же, собственно, представляет собой ген.
В типичном эксперименте по составлению
карты штамм В бактерий заражают двумя различными мутантами rII и получают потомство, состоящее в основном из тех же
двух типов мутантов, как и родители, но и, кроме того, из нескольких
рекомбинантов. Общее число фагов определяется в результате подсчета стерильных
пятен на штамме В. Если выращивать потомство на штамме бактерий К, то мутантные
типы вымирают и остаются только рекомбинантные, так что появляется возможность
установить более точное их соотношение1. Бензер доказал, что
рекомбинации происходят в основном между аллелями внутри локуса rII, и смог определить генетическое расстояние между каждыми двумя
мутантными участками (сайтами) и даже составить карту этих аллелей. Небольшая
часть этой карты выглядит следующим образом:

208
210
Рис. 8.2. С помощью
комплемвнтационного теста можно определить, происходят ли две мутации внутри
одного гена или нет.
Бактерии одновременно заражают
двумя фагами с двумя различными мутациями, которые затрагивают либо один ген (слева), либо два гена (справа). Если
мутации затрагивают один ген, то ни в одном фаге не создается нормальной копии
гена, поэтому фаги не могут размножаться. Но если мутации затрагивают оба гена,
то один фаг имеет нормальный ген А, а другой - нормальный ген В, и
оба гена дополняют друг друга. Обратите внимание, что этот тест не имеет ничего
общего с кроссинговером
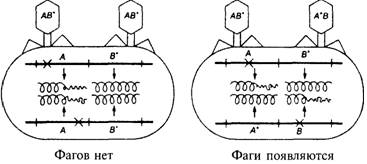
группы. Ни один из мутантов по левой
группе не дополнял мутантов по этой же группе, и то же самое было с правой
частью. В то же время любой мутант из левой группы оказывался комплементарным к
любому мутанту из правой группы. Эти результаты доказывают, что область rII действительно включает в себя два гена. (Хотя Бензер
называл отдельную функциональную единицу цистроном,
сейчас цистроном называют то же, что и ген.) Комплементационные
тесты, подобные этому, в наши дни применяют ко всем организмам, чтобы узнать,
происходят ли две мутации внутри одного гена или нет, и определить таким
образом границу между генами.
211
Что же такое ген?
Вернемся к определению гена. В
классической генетике словом 'ген' обозначалась единица генетического
материала, выделяемая по трем критериям: по функции, мутации и рекомбинации.
Изначально предполагалось, что ген - это функциональная единица, то есть нечто,
определяющее отдельный признак. Такое представление сохранилось и до сих пор,
но сейчас нам известно, что на один и тот же признак могут воздействовать
различные гены и что при мутации гены могут давать один и тот же фенотип. Кроме
того, ген определяли как единицу мутации. Эксперименты Бензера показали, что
ген представляет собой линейную последовательность многих участков, в которых
возможны разные мутации, и мы только что показали, как в комплементационных
тестах можно выделять гены на основе происходящих в них мутаций. При этом ген
понимается как последовательность, кодирующая синтез отдельной полипептидной
цепи, и это представление основано на концепции Бидла и Тэйтема 'один ген - один
фермент'. Гены они определяли и как единицы рекомбинаций, хотя сейчас известно,
что гены не представляют собой неделимые 'бусины' на цепи, а рекомбинации
происходят и внутри генов. Это и следовало ожидать, если предположить, что ген
представляет собой всего лишь участок ДНК, любые нуклеотидные пары которой
могут изменяться, в результате мутации и рекомбинаций.
В свете последних исследований, особенно секвенирования (определения последовательности
ДНК), приходится по-новому подходить к вопросу о том,
212
Бактерии и фаги, которые их атакуют,
находятся в состоянии непрерывной химической войны. Бактерии, оказывающие
сопротивление фаговой инфекции, получают преимущество в борьбе за существование,
и они выживают с большей вероятностью. Точно так же фаги, преодолевающие
защитные барьеры бактерий, получают определенное преимущество. Бактерии
производят рестрикционные
214
ферменты - эндонуклеазы
(рестриктазы), которые
атакуют молекулы ДНК, разрезая их фосфодиэфирные связи (эндо- означает, что
они разрезают молекулу изнутри, а не по краям). Эти ферменты образуют рестрикционную систему, которая разрушает
фаговые ДНК. Сейчас разработаны простые и быстрые методы определения
последовательности молекул ДНК. Секвенирование ДНК показывает, что каждая
эндонкулеаза очень специфична и что она разрезает только очень короткую
последовательность ДНК, чаще всего так называемый палиндром.
Палиндром - это последовательность букв, которая одинаково читается как
обычным способом, так и задом наперед подобно известным фразам: 'А роза упала
на лапу Азора' или 'А кит на море - романтика!' Молекулярный палиндром - это
последовательность оснований, которая также читается одинаково в любом
направлении, например:
3'-GAATTC-5' или 5'-CTTAAG-3'.
Фермент, атакующий именно эту
последовательность, синтезируется штаммом Е. coli RY13, так что если фаг с такой последовательностью
попытается атаковать бактерию, фермент разрежет его ДНК на фрагменты и
остановит инфекцию. На рис. 8.3 указаны последовательности, которые
подвергаются атаке со стороны ферментов, выделенных из различных типов
бактерий. (Источник каждого фермента обозначен трехбуквенным сокращением по
названию бактерии, например, фермент, выделенный из Е.
coli RY13, называется
EcoRI.)
Почему в таком случае бактерии не
разрушают собственную ДНК? В них параллельно рестрикционной включается модификационная система, фер-
215
менты которой добавляют метиловую группу (СН3)
к аденинам последовательности, блокируя тем самым действие рестрикционной
эндонуклеазы. Некоторые фаги добавляют метиловую группу к своей ДНК и
становятся невосприимчивыми к такой эндонуклеазе.
Рис. 8.3. Молекулы ДНК можно легко
разделить

216
218
следовательность прочитывают по
распределению полос.
Сейчас известно и доступно для применения
множество типов рестрикционных ферментов. Они разрезают ДНК на различные
последовательности, и их можно использовать для анализа структуры ДНК и
составления хромосомных карт методом рестрикционного картирования. На
рестрикционной карте отмечено относительное расположение участков (сайтов
рестрикции), которые
вырезают различные нуклеазы. С помощью других методов можно сопоставить эту
карту с генетической картой. В данном случае применяют технологию
электрофореза, описанную во вставке (с. 216-218), с помощью которой разделяют
ДНК на фрагменты и определяют их относительный размер. Рестрикционное
картирование лучше объяснить на примере. У многих вирусов животных имеются
маленькие кольцевые ДНК. Предположим, мы выделили ДНК вируса длиной в четыре
килобазы (kb) и порезали ее ферментом ЕсоRI на фрагменты.
Пропустив их через гель, мы определили их длину: 0,4; 0,8; 1,3 и 1,5 kb. Это значит, что в геноме находится четыре участка
рестрикции EcoRI,
которые могут располагаться
по-разному.
Порежем вирусную ДНК снова при помощи EcoRI, но на этот раз уменьшим время обработки ДНК ферментом, чтобы некоторые
ДНК были порезаны не полностью. Наряду с прежними четырьмя фрагментами получаем
новые фрагменты длиной 1,7;
219
222
полиморфизм служит нейтральным
гетерозиготным маркером, с помощью которого можно определять близлежащие гены,
особенно при составлении карт маркеров, приводящих к разным фенотипам. Кроме
того, если RFLP расположен близко от дефектного аллеля, то его можно
использовать для обнаружения этого аллеля подобно участку рестрикции внутри
гена гемоглобина. В геноме встречаются и другие типы нейтральных вариаций
последовательности, и RFLP оказался первым среди открытых учеными. Все они могут
быть использованы для составления карт и обнаружения рецессивных дефектных
аллелей.
Глава 9. РАСШИФРОВКА КОДА ЖИЗНИ
Как только Уотсон и Крик предложили свою
модель ДНК, ученые поняли, что линейная последовательность оснований ДНК
составляет ряд ключевых слов, или кодонов, соответствующих
линейной последовательности аминокислот в белках. Кроме того, как заметил Крик,
поскольку и ДНК, и белки представляют собой линейные последовательности
элементов, обе последовательности должны быть колинеарными.
Это значит, что первый кодон гена должен кодировать синтез первой
аминокислоты, второй ген - второй аминокислоты и т. д. Оставалось только
логически выяснить, какие именно сочетания четырех оснований A, G, С и Т
образуют эти кодоны.
Белки состоят из 20 видов аминокислот.
Предположим, что кодону соответствуют последовательности из двух оснований,
например АА или СТ. Так как оснований всего четыре, получается: 4 х 4 = 16
сочетаний. Этого недостаточно для 20 аминокислот. Далее предположим, что кодон
- это триплет, то есть последовательность из трех оснований. Теперь получается:
4 х 4 х 4 = 64 сочетания, то есть больше 20. В таком случае либо 44 триплета
являются бессмысленными, либо мы имеем
224
Как строятся белки!
Итак, информация, определяющая порядок
аминокислот в белке, хранится в ДНК в виде ряда триплетных кодонов. Но как
последовательность оснований ДНК превращается в реальный продукт? Конечно,
чертежи и схемы для строительства очень важны, но для построения здания требуется
ряд сложных операций.
Строение клеток эукариот накладывает на
такие операции некоторые ограничения. ДНК хранится в хромосомах,
располагающихся в ядре, а белки синтезируются в цитоплазме, в органеллах - рибосомах, которые по большей части покрывают
мембраны эндоплазматической сети (рис. 9.1). Если ДНК содержит чертежи для
строительства белков, а сама 'стройка' располагается в цитоплазме, то как
чертежи попадают на стройку? Это все равно, что по-
228
Рис. 9.1. Эндоплазматическая сеть
клеток эукариот состоит из мембран, обычно расположенных параллельно друг другу
и покрытых крохотными частицами - рибосомами, которые служат фабриками по
производству белка

лучить важную формулу из редкой книги,
которую не позволяют выносить из библиотеки. В этом случае нужно сделать
фотокопию и принести фотокопию в мастерскую. Точно так же обстоит дело и в
клетке. Копией служит РНК.
Молекулы РНК: инструменты для синтеза белка
В 1940-х годах, когда ученые еще
недостаточно хорошо представляли строение нуклеиновых кислот, были получены
доказательства того, что син-
229
РНК-транскрипция
Сейчас доказано, что РНК образуется в
результате того же спаривания комплементарных оснований, с помощью которого
образуется и двойная спираль ДНК из одинарной цепи (рис. 9.2). Этот процесс
называется транскрипцией. Его
выполняет особый сложный фермент - РНК-полимераза.
Возле каждого гена располагается участок - промотор с последовательностью оснований, к
которым при-
Рис. 9.2. В процессе транскрипции
на одной из цепей ДНК образуется комплементарная ей цепь РНК.
Этот процесс похож на
синтез новой цепи ДНК во время репликации, но со следующими отличиями: а) копируется только одна цепь ДНК; б) синтезируется РНК, а не ДНК; в) процесс происходит при помощи фермента РНК-полимеразы
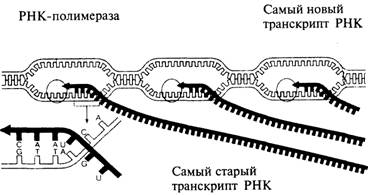
233
234
Рис. 9.3. Электронная
микрофотография транскрипции РНК. ДНК имеет вид тонкой нити, проходящей через
центр каждого участка, похожего по форме на перо.
От ДНК отходят молекулы
РНК: транскрипция самых длинных уже почти закончилась, транскрипция самых коротких
только началась. Это рибосомные РНК, которые входят в структуру рибосом
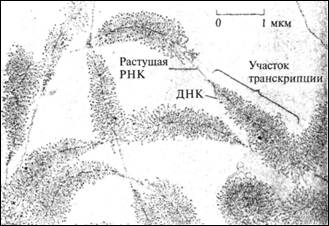
дения, или 'отжига', добавить
одноцепочечные молекулы РНК, то образуются также и гибридные молекулы ДНК-РНК,
но для этого требуются молекулы, полностью комплементарные цепям ДНК. Обычно в
ходе таких экспериментов денатурируют ДНК и пропускают раствор одинарных цепей
через нитроцеллюлозные фильтры. Если на эти фильтры нанести раствор меченой
РНК, то РНК-транскрипты с комплементарными последовательностями соединятся с
ДНК, и их можно будет легко обнаружить, измерив радиоактивность фильтра.
Молекулы РНК, не имеющие комплементарной последовательности, не соединятся с
ДНК и пройдут через фильтр.
235
236
ток, составляющие часть аппарата по
синтезу белка, как будет показано далее.
Перенос информации с ДНК на РНК
называется транскрипцией, а перенос
этой информации с мРНК в белок - трансляцией. Обычно
матричные РНК в течение некоторого времени программируют рибосомы на производство
определенного белка, а затем разрушаются. Рибосома сама устанавливает рамку
считывания, пропуская мРНК между своими половинами и передвигая мРНК по три
основания за раз, начиная с 5'-конца и заканчивая 3'-концом. Одна молекула мРНК
может проходить одновременно через несколько рибосом, которые синтезируют один
вид белка друг за другом. Несколько рибосом, присоединенных к одной мРНК,
называются полирибосомой.
Каждая мРНК содержит серию кодонов,
скопированных с ДНК, но в аминокислотах нет химических структур, которые
распознавали бы нужные кодоны. Тут в действие вступают транспортные РНК (рис.
9.4). В каждой клетке имеется по меньшей мере по одному виду тРНК для каждой из
20 аминокислот. В клетке также содержится 20 разных видов ферментов (аминоацил-РНК-синтетаз),
по одному на каждую аминокислоту. Каждый фермент распознает строго определенную
тРНК вместе с соответствующей ей аминокислотой и соединяет их. В результате
получается аминоацил-тРНК. Три основания тРНК образуют антикодон, комплементарный кодону мРНК, так
что каждая аминоацил-тРНК может подсоединяться к мРНК в нужном месте.
237
Рис. 9.4. Общая структура молекулы
транспортной РНК.
Обратите внимание на
значительные внутренние сгибы, образованные парами G-C и A-U. Аминокислота присоединяется к одному концу,
а на другом конце располагается петля с антикодоном, который распознает кодон
матричной РНК. Структуры некоторых оснований слегка изменены добавлением
небольших химических групп, таких как гидроксильная и метильная группы
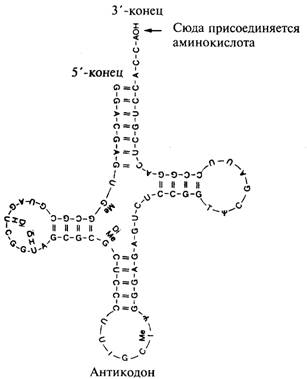
238
242
жет подвергаться мутациям и рекомбинациям
без выраженного эффекта. (О повторяющейся ДНК в геноме человека говорится в гл.
12.)
К 1962 году благодаря работам Крика и его
коллег, о которых говорилось ранее, было установлено, что генетический код
состоит из триплетов. После этого перед исследователями встала другая непростая
задача: определить, какие именно аминокислоты кодирует тот или иной триплет.
Как часто бывает, открытие было сделано почти случайно, после чего весь код был
расшифрован за несколько лет - одно из величайших достижений молекулярной
биологии! В 1961 году Маршалл Ниренберг и Филипп Ледер разрабатывали методы
искусственного синтеза белка, смешивая рибосомы, источники энергии,
активирующие ферменты, тРНК и другие компоненты. В одну из контрольных смесей,
синтез белка в которой не ожидался, они добавили искусственную РНК, состоявшую
исключительно из урацила, то есть полимера с нуклеотидной последовательностью U-U-U-U-U-, называемого полиуридиловой
кислотой. Вопреки ожиданиям эта кислота повела себя, как
информационная РНК, и стимулировала синтез белка. В такой среде с
полиуридиловой кислотой синтезировался только полифенил-аланин, то есть
последовательность U-U-U должна была
кодировать производство одной аминокислоты, а именно фенилаланина.
После этого открытия началось настоящее
состязание между лабораториями Ниренберга и Се-
243
веро Очоа, в которых с помощью
синтетических РНК старались подобрать код к каждой аминокислоте. Поскольку
фермент, создающий такие синтетические молекулы, соединяет основания в
случайной последовательности, поначалу приходилось полагаться на статистический
анализ получающихся полипептидов. Настоящий прорыв был сделан только тогда,
когда Ниренберг и Генрих Матей попытались синтезировать мини-мРНК с тремя
нуклеотидами в известной последовательности. Обнаружилось, что в искусственной
среде каждый из этих триплетов присоединялся к рибосоме и распознавался только
одним видом тРНК. Исходя из этого легко было узнать, какие аминокислоты
кодировались тем или иным триплетом. Исследователи выяснили, что UUU и UUC (если читать их в направлении 5'-> 3'), например,
присоединяют к себе тРНК фенилаланина, GUU - тРНК валина, UUG - тРНК лейцина, a UGU присоединяет
тРНК цистеина. В конце концов с помощью ученых из других лабораторий был
расшифрован генетический код всех аминокислот и получен своеобразный
'генетический словарь' (табл. 9.1).
На основании приведенной таблицы можно
сделать ряд выводов. Как и предсказывал Крик, код оказался вырожденным, но при
этом количество кодонов, определяющих ту или иную аминокислоту, варьируется от
одного (метионин, триптофан) до шести (лейцин, серии, аргинин). Кроме того,
вырожденность кода довольно регулярна. В любом случае весь смысл определяют два
первых основания (в направлении 5' -> 3'). В восьми случаях не имеет
значения, какое за ними следует третье основание, так как аминокислота
определяется и без
244
Таблица 9.1. Генетический код
Примечание. Каждый из 64
триплетов либо кодирует одну из аминокислот (обозначенных трехбуквенными
сокращениями), либо означает конец синтеза полипептидной цепи.
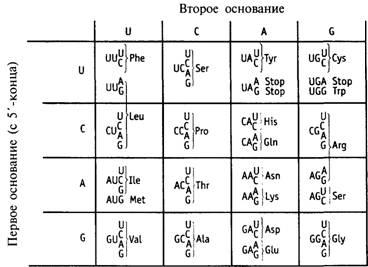
него. В 12 случаях смысл определяет выбор
между пурином (A, G) или пиримидином (U, С).
Триплет AUG, кодирующий метионин, в начале гена почти всегда
используется для специальной тРНК, переносящей метионин с блокированной
аминогруппой (N-формилметионин). В другие места
белка метионин переносит другая тРНК.
245
Гипотезу о колинеарности гена белку можно
было подтвердить, показав, что последовательность мутаций гена соответствует
изменениям последовательности аминокислот, к которым приводят эти мутации. Для
этого необходим ген с известной картой, кодирующий белок, последовательность
которого также известна. Чарльз Янофски в этих целях исследовал гены ферментов,
синтезирующие триптофан (trp-гены)
в Е. coli. Янофски с коллегами
использовал ген trpA, кодирующий
производство белка А, часть фермента триптофансинтетазы. Исследователи
определили полную последовательность из 267 аминокислот в белке А дикого типа и
в 10 белках-мутантах по локусу trpA. Каждая мутация заменяла во всем белке только одну аминокислоту. Они
составили также карты мутационных участков для 10 мутантов и показали, что
последовательность мутационных участков и последовательность заменяемых
аминокислот в мутантных белках соответствуют друг другу (рис. 9.6).
Генетическое расстояние между двумя участками мутаций пропорционально расстоянию
между аминокислотами, на которые они воздействуют, так что генетическое
расстояние, определяемое в результате рекомбинаций, действительно соответствует
физическим расстояниям внутри гена. Обратите внимание на то, что в двух случаях
два аллеля, выделяемые на основе рекомбинаций, соответствуют разным заменам
одной и той же аминокислоты. Обычный глициновый остаток в позиции 210 у мутанта
Л23 заменяется на глутамин, а у
мутанта Л46 - на глутаминовую
кислоту. Согласно генетическому коду глицину соответствует кодон GGX
246
Рис. 9.6. Колинеарность гена и
синтезируемого им белка.
Последовательность
мутационных участков в ДНК идентична последовательности аминокислот белка. В
двух случаях соседние мутации определяют разные замены одной и той же
аминокислоты
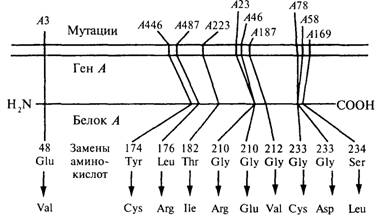
(где X обозначает А
или G), и у мутанта А23 он
заменяется на кодон CGX, а у мутанта А46 - на GAX. Мутации А78 и
А58 также по-разному затрагивают один
и тот же кодон. Таким образом, ген состоит из линейной последовательности
кодонов, определяющей последовательность аминокислот, и кодирующий элемент
каждой аминокислоты может в результате рекомбинаций разделяться на части.
Этот впечатляющий генетический
эксперимент был подтвержден другим замечательным экспериментом Джорджа
Стейзингера и его коллег. Они получили мутанты фага Т4, производящие мутантный
фермент лизоцим, и выделили синтезируемые ими белки, определив
последовательность аминокислот. Под воздействием профлавина они вызвали
247
мутации этого гена и получили супрессор.
Белок двойного мутанта оказался почти дикого типа, за исключением небольшого
бессмысленного набора аминокислот, соответствующих бессмысленному набору
оснований между двумя мутационными участками. Это было изящное и логичное
продолжение работы Крика.
Все описанные генетические опыты помогли
ученым разработать модель работы гена. Они проводились независимо от
биохимических исследований и вместе с тем подтвердили, что код ДНК
транслируется в структуру белка.
Три кодона из 64 не служат кодом для
аминокислоты. Они означают конец синтеза белка и называются терминирующими или нонсенс-(стоп-) кодонами (stop codon). Их
существование было подтверждено нонсенс-мутациями,
которые приводили к образованию именно таких кодонов. Обычно эти
триплеты располагаются в конце гена, но если в результате мутации один из
терминирующих кодонов оказывается внутри гена, то синтез белка немедленно
прерывается. Одна из серий нонсенс-мутаций, а именно амбер-мутации, по определению имеет дефекты, супрессором
которых может быть другая мутация, затрагивающая механизм синтеза белка. В
результате супрессорной мутации образуется тРНК, способная распознавать
нонсенс-кодон и вставлять в полипептидную цепь аминокислоту, делая тем самым
этот кодон смысловым. Секвенирование нормальных белков показало, что амбер-
248
Глава 10. НАСЛЕДСТВЕННОСТЬ В МИРЕ БАКТЕРИЙ
Представители классической генетики едва
ли смели мечтать о тех возможностях, какие открываются перед современными
учеными, проводящими эксперименты на бактериях и вирусах бактерий. В этой главе
мы познакомимся с некоторыми результатами их работ и принципами, согласно
которым функционирует генетический аппарат не только бактерий, но и всех живых
организмов. Мы узнаем также о перспективах современной генетики и получим
представление о вопросах, какие она ставит перед человечеством. Способы
клонирования ДНК, воспроизводства ее последовательности, а также создания
генетически модифицированных организмов были впервые опробованы в исследованиях
по генетике бактерий и вирусов.
Разные виды бактерий можно различать по
фенотипическим признакам, таким как форма, цвет и другие характерные
подробности их колоний. Но большой прогресс в генетике бактерий был достигнут в
ходе исследований мутантов одного вида, пре-
251
257
Рис. 10.2. Карта хромосомы Е.
coli.
Единицами служат минуты,
то есть промежутки времени, через которые переносятся гены при скрещивании Hfr х F-. Отдельные участки карты с большим
количеством известных генов более подробно показаны на внешних сегментах. Более
мелкие подробности определяются в процессе трансдукции, о чем говорится далее.
Данная карта была опубликована несколько лет назад, и сейчас существуют более
подробные карты, которые заняли бы несколько страниц
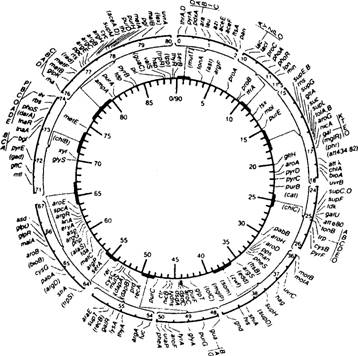
жаться различные гены, однако в отличие
от вирусов плазмиды не образуют внеклеточных частиц (вирионов) ни на одной из
стадий своего развития. Многие плазмиды могут переносить свои копии в
258
другие клетки подобно F, но некоторые плазмиды просто располагаются в
клетках-хозяевах и не могут самостоятельно переходить из клетки в клетку.
Фактор F - это пример подкласса плазмид - эписом, которые могут существовать в клетках
хозяев как отдельные элементы или встраиваться в хромосомы хозяев.
Факторы резистентности и устойчивость
к антибиотикам
В 1955 году одна жительница Японии
вернулась из Гонконга с разновидностью дизентерии, вызываемой бактерией рода Shigella. Инфекцию Shigella легко
лечить антибиотиками, но эти бактерии оказались устойчивыми сразу к четырем
разным антибиотикам: сульфаниламиду, стрептомицину, хлорамфениколу и
тетрациклину. Это казалось очень подозрительным, потому что обычно устойчивость
к антибиотикам встречается редко. Если бактерии поместить в среду с
пенициллином, то устойчивой к нему окажется примерно одна на 107
клеток. Точно так же выживет и одна из 107 клеток в среде
со стрептомицином. Если же бактерии поместить в среду с двумя антибиотиками, то
можно ожидать, что выживет одна из 1014 клеток. Поэтому
одновременная устойчивость к четырем видам химических веществ выглядит почти
чудом. Но все же за последующие годы бактерии дизентерии в Японии стали очень
устойчивыми к антибиотикам, и количество случаев множественной резистентности
многократно возросло.
259
264
Биологи, проводившие эксперименты с
фагами до Второй мировой войны, часто утверждали, что некоторые штаммы бактерий
переносят вирусы, которые иногда непредсказуемо проявляют себя в растущих
культурах. Ученые школы Дельбрюка не воспринимали серьезно подобные утверждения
и отвергали эти факты, полагая, что бактерии заражались из-за недостаточно
стерильных методов работы. Однако в 1950 году Андре Львов и Антуанетта Гутман
из Парижа доказали существование явления, названного лизогенией, и содержание неактивных форм фагов, называемых
профагами, в некоторых бактериях. Лизогенные штаммы имеют иммунитет против тех
фагов, переносчиками которых они являются. Иногда профаг в лизогенной клетке
спонтанно индуцируется, то есть
переходит в активное, или литическое, состояние.
Его ДНК быстро размножается, и клетка наполняется частицами фага, после чего
лизирует, то есть разрушается, как если бы она была заражена фагом наподобие
Т4. Львов позже обнаружил, что можно индуцировать литическую фазу, подвергнув
бактерии воздействию ультрафиолетового света или других реагентов. Так
исследователи смогли контролировать рост фагов.
Фаги, существующие в лизогенных
бактериях, называются умеренными, в
противоположность вирулентным фагам,
таким как Т2 или Т4, которые всегда быстро размножаются и убивают своего
хозяина. Когда умеренный фаг заражает чувствительную (нелизогенную) клетку, он либо размножается
литически, либо становится профагом и преобразует бактерию в лизогенную клетку.
Из этой клетки
265
267
Этот кусок ДНК может быть встроен в
фаговую частицу, и тогда фаг лямбда станет переносить гены gal или bio в
другие клетки. Это так называемая специфическая трансдукция,
которая затрагивает лишь некоторые отдельные гены. Другие фаги, подобные P1, могут осуществлять общую
трансдукцию и переносить любые гены. Во время размножения такого
фага хромосома бактерии режется на мелкие куски, и иногда один из кусков
попадает в головку фага вместо его генома. Затем такой фаг может присоединиться
к другой клетке и внедрить в нее содержимое своей оболочки, то есть
бактериальные гены. Трансдукция послужила эффективным средством для определения
тонкой структуры бактериальных генов и составления карт бактериальных хромосом.
Фаги переносят только небольшие кусочки ДНК, и анализ того, каким образом эти
кусочки рекомбинируют с новой хромосомой хозяина, позволяет установить
последовательность бактериальных маркеров.
В 1955 году Джошуа Ледерберг предположил,
что трансдуцирующие вирусы можно использовать для введения генов в клетки
человека. В то время такая идея казалась чистой фантазией, но сейчас она все
более и более приобретает черты реальности, особенно в свете развития
технологии рекомбинантных ДНК, описанной в гл. 12.
Трансдукцию можно использовать для того,
чтобы: 1) внедрить доминантные 'хорошие' гены и исправить наследственный
дефектный фенотип;
268
2) внедрить гены в пищевые растения и
увеличить их полезность; 3) изменить бактерии так, чтобы они выполняли полезные
биологические функции; 4) вызывать заболевание в определенном организме.
Главный технический вопрос заключается в том, могут ли вирусы захватывать гены
животных и внедрять их в другие клетки. Ответ на этот вопрос положительный.
Например, если вирусом простого герпеса заразить штамм мышиных клеток, в
которых отсутствует определенный белок (тимидинкиназа), то у 0,1% клеток
появляется нормальный аллель. Нормальный аллель могли привнести только вирусы.
Клетки с привнесенным аллелем довольно стабильны и производят тимидинкиназу на
протяжении 8 месяцев. В других экспериментах используют аденовирусы и другие
вирусы животных, с помощью которых переносят гены, например для лечения
генетических заболеваний. Были выведены трансдуцирующие полиомовирусы, которые
могут захватывать гены мыши и переносить их в ядро клетки человеческого
эмбриона.
Некоторые медицинские технологии иногда
разрабатываются до того, как удается определить этические правила их
применения. В истории науки так бывало часто, например когда Бантинг и Бест разрабатывали
и тестировали опытные образцы инсулина или когда Кристиан Барнард поразил
общественность заявлением об удачной трансплантации сердца. То же можно сказать
и о трансдукции генов в клетки человека. В 1958 году Стэнли Роджерс изучал
вирус папилломы Шопа, которая вызывает повреждения кожи у кроликов. Он
обнаружил у инфицированных клеток высокий уровень содержания фермента аргиназы,
вызванный, по всей ви-
269
Глава 11. РЕГУЛЯЦИЯ ГЕНОВ И РАЗВИТИЕ ОРГАНИЗМА
По мере чтения книги, как и на протяжении
всей истории генетики, наше представление о генах постоянно изменялось. Если
сначала мы считали ген неопределенным фактором, который каким-то образом
передается по наследству, то теперь мы называем геном специфическую
последовательность ДНК (иногда РНК), которая определяет структуру белка. Всякий
ген располагается в хромосоме; все гены в своей совокупности составляют геном,
определяющий жизнедеятельность организма. Но в этой картине ясно еще не все. Вы
можете сказать: 'Ну хорошо, я понимаю, как устроены гены и как они передают
инструкции по синтезу самых различных ферментов и белков. Но посмотрите на
меня! Я же не просто мешок с ферментами и даже не множество маленьких мешочков
с ферментами, присоединенных к костям. Если мой геном и вправду определяет мою
жизнедеятельность на протяжении всех этих лет, то они должны были как-то
сделать так, чтобы я рос и из единственной клетки-зиготы превратился в
высокоорганизованную структуру из многих видов клеток. Я хочу узнать, как мои
гены определили мой рост и мое развитие'. Если вам в
274
голову приходили подобные мысли, то вы
вступили на тот же путь, что и многие современные генетики, посвятившие свою
профессиональную жизнь поиску ответов на вопросы о механизме развития организмов.
Здесь важно обратить внимание на то, что
из одной клетки развиваются различные виды клеток. Во многих книгах по биологии
и анатомии описаны различные типы тканей, из которых состоит тело человека,
животных и растений. Все эти ткани, в свою очередь, состоят из клеток разных
типов. Большинство наших клеток представляют собой подобие кубиков или
цилиндров, которые образуют большие органы вроде печени, трубы вроде пищевода и
кровяных сосудов, а также нижнюю часть кожного покрова. Плоские клетки создают
гладкие поверхности внутри сосудов или внешние оболочки. Клетки мышц
представляют собой либо очень длинные цилиндры, либо небольшие веретена с
упорядоченными белковыми волокнами, которые вытягиваются и сокращаются. Наша
нервная система содержит клетки с длинными и тонкими отростками (некоторые даже
более метра в длину), которые очень быстро передают сигналы по всему телу.
Всего в нашем организме более сотни различных видов клеток, и чем больше мы их
исследуем, тем больше узнаем об их специализации.
Таким образом, можно перефразировать наш
вопрос: 'Как все эти клетки стали отличаться друг от друга?' Мы уже знаем
достаточно, чтобы сказать, что они отличаются прежде всего тем, что производят
разные виды белков. Если клетка вырабатывает белок, кодируемый определенным
геном, то говорят, что этот ген выражен или
что происходит
275
290
Рис. 11.3. Начальные взаимодействия
генов, определяющих пространственную ориентацию эмбриона плодовой мушки.
В передний конец
внедряется мРНК bicoid и синтезируемый на ее основе белок Bicoid включает транскрипцию гена hunchback. Это приводит к транскрипции генов
головы и груди. В задний конец внедряется мРНК nanos и синтезируемый ею белок. Nanos
подавляет экспрессию
белка Hunchback. Как следствие включаются гены брюшка
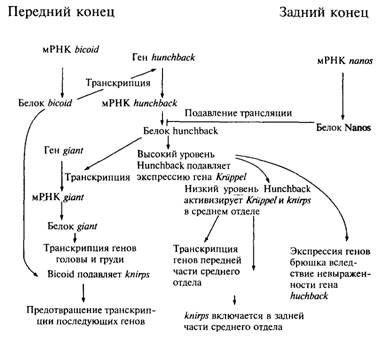
Решающим событием служит внедрение мРНК bicoid в тот конец, который позже станет передним (и где
будет расположена голова), а также внедрение мРНК nanos в тот конец, который позже станет задним. Белки,
транслируемые с каждой из этих мРНК, либо способствуют синтезу хотя бы одного
291
из белков, либо подавляют синтез. Затем
цепь синтеза белков идет по нарастающей, пока белки не образуют разные
структуры.
Как только начальное направление
определено, включаются другие гены, которые делят эмбрион на сегменты,
поскольку тело мухи членится на отдельные сегменты: 5 сегментов в области
головы, 3 грудных сегмента и 11 в области брюшка. Задействованные при этом
белки можно наблюдать при помощи подкрашивания их антител. Если развивающийся
эмбрион погрузить в раствор антител, то антитела сконцентрируются там, где
расположены соответствующие белки. Такие опыты показали, что сначала включается
серия генов gap. Кодируемые ими белки
активизируют серию генов pair-rule, которые делят эмбрион на 14 сегментов. Затем включаются гены segment-polarity, которые делят каждый сегмент на переднюю и заднюю части. После этого
начинает действовать серия генов homeotic, которые определяют строение каждого сегмента с присущими ему
признаками. Именно эти гены первыми были опознаны в мутантах необычной формы.
Например, мутанты Antennapedia имеют на голове вместо усиков пару ног. Спаривая между
собой различных гомеотических мутантов, Эд Льюис вместо обычных двукрылых
вырастил четырехкрылых мушек.
Одна из самых интересных серий событий с
участием нескольких генов происходит при формировании глаза мушки дрозофилы.
Сложный глаз насекомого состоит приблизительно из 800 элементов. Отдельный
элемент, омматидий, состоит пример-
292
но из 20 клеток, расположенных в
определенном порядке. Эти клетки образуют тонкий слой и взаимодействуют только
со своими соседями; выше и ниже клеток нет. Обычно эти клетки становятся
клетками хрусталика, но если они вступают в серию генетически
запрограммированных взаимодействий друг с другом, то они специализируются в
фоторецепторы, которые воспринимают свет и посылают в мозг сигналы.
Дифференциация в каждом омматидий происходит в строго определенной
последовательности (рис. 11.4). Сначала специализируется центральный
фоторецептор, R8. Далее он вызывает
специализацию своих соседей, R2 и R5. Потом они, в свою очередь, вызывают специализацию
своих соседей с одной стороны, R3 и R4, и с другой стороны, R1 и R6. Под конец
происходит специализация R7. Оставшиеся
клетки группы становятся хрусталиком и другими структурами.
Дифференциация начинается после того, как
через слой клеток из передней в заднюю часть проходит волна, видимая, как
заметное углубление. После прохождения волны клетки начинают делиться и затем
останавливаются на ранней стадии клеточного цикла. Ориентация волны, а
следовательно, и группы фоторецепторов, зависит от экспрессии гена wingless (wg), продуктом которого
является белок, сигнализирующий о начале выработки серии белков, выполняющих
специфические действия по дифференциации. Клетки за волной начинают
дифференцироваться и выделять белок Hedgehog (Hh). Hh растворяется
в недифференцированных клетках и стимулирует их производить белок Decapentaplegic (Dpp). Dpp растворяется
в еще большем количестве передних клеток, заставляя их дифференцироваться и
293
Рис. 11.4. Восемь клеток, которые
становятся фоторецепторами в омматидии глаза дрозофилы, специализируются в
определенном порядке, который зависит от взаимодействия нескольких генов и
кодируемых ими белков
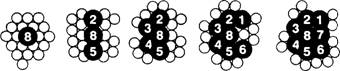
выделять Hh, который в свою очередь запускает очередной цикл
производства Dpp. Так циклическое производство
белков Hh и Dpp заставляет
волну двигаться вперед.
Чтобы понять, как специализируются
следующие клетки, рассмотрим клетку R7, хотя
она и специализируется в последнюю очередь. Мутация sevenless (sev) происходит в гене,
который требуется для дифференциации R7. Клетка
R7 отличается тем, что это рецептор ультрафиолетового
света, и мутанты sevenless могут
быть опознаны по тому, что они не воспринимают ультрафиолетовый свет. Белок Sevenless служит
рецептором на поверхности клетки, которая станет R7, и, как лиганд, он связывается с белком Boss, который кодируется геном bride-of-sevenless (boss) и вырабатывается
клеткой R8. Таким образом, белок Boss клетки R8 дает инструкции соседней клетке по превращению в
специализированную клетку R7.
Мы еще не знаем до конца, как именно
запрограммированы все эти взаимодействия, но анализ некоторых мутантов вывил
общую картину: одна из
294
клеток вырабатывает определенные белки,
которые взаимодействуют с белками соседних клеток, включая один набор генов и
выключая другой. Если вернуться к вопросу, поставленному в начале главы, то
придется ответить, что существуют разные механизмы развития. Одни гены кодируют
белки, которые непосредственно связываются с другими генами, включая их или
выключая. Другой класс генов кодирует белки, регулирующие трансляцию
специфических мРНК в соответствующие им белки. Некоторые гены кодируют синтез
белков, переходящих из одной клетки в другую и заставляющих их меняться или
противодействующих переменам. Есть и такие гены, которые кодируют белки,
взаимодействующие с другими белками на поверхности близлежащих клеток. Развитие
эмбриона зависит от сложного переплетения разнообразных видов взаимодействий,
среди которых мы упомянули лишь несколько самых распространенных.
Глава 12. ВМЕШАТЕЛЬСТВО В СТРОЕНИЕ ДНК: ВОЗВРАЩЕНИЕ ЭПИМЕТЕЯ?
В древнегреческих мифах говорилось о
титанах - расе гигантов, рожденных богами прежде расы людей. Титану Эпиметею
боги поручили создать животных и растения, распределив между ними разнообразные
свойства. Но Эпиметеи (его имя переводится как 'думающий после') взялся за дело
слишком поспешно и исчерпал весь запас свойств, прежде чем дошел до людей.
Поэтому он попросил своего брата Прометея похитить у богов огонь и подарить его
людям. Этот поступок навлек гнев Зевса, пославшего Эпиметею прекрасную жену
Пандору, которая должна была доставить ему немало страданий. Пандора открыла
запретный ящик (также посланный на землю Зевсом), в котором хранились все
бедствия и несчастья мира. Сегодня многие уверены в том, что генетики подобно
современным Эпиметеям бездумно вмешиваются в тайны природы и могут нечаянно
выпустить разнообразные бедствия из научного ящика Пандоры. Эти страхи еще
более усилились, после того как генетики научились изменять строение ДНК.
Теперь они стали подобны богам, получив возможность создавать организмы,
которые никогда не су-
296
шествовали и не могли бы существовать без
этой новой технологии.
Мы уже говорили, как начиная с ранних
эпох скотоводства и земледелия люди пытались выращивать новые сорта растений и
разводить новые породы животных. Законы Менделя в ХХ веке помогли людям
улучшить традиционные способы селекции, но все изменения до сих пор происходили
только внутри отдельных видов. С
появлением генных технологий в одном организме можно объединить признаки сразу
нескольких других организмов, подобно тому как в древнегреческой мифологии
созданное богами огнедышащее чудовище Химера изображалось с телом льва, головой
козла и хвостом змеи. В данной главе мы поговорим именно об этом виде
вмешательства в ДНК. Затрагиваемые при этом этические и социальные вопросы
обсуждаются в следующей главе.
К 1972 году Анни Чанг, Поль Берг и Сеймур
Коэн установили, что при помощи рестрикционных ферментов, рестриктаз, можно порезать две любые молекулы
ДНК и сделать из них одну рекомбинантную ДНК.
Именно это открытие легло в основу новой технологии, революционным образом
преобразившей современную генетику и молекулярную биологию. Метод основан на
стремлении комплементарных одинарных цепей нуклеиновых кислот соединиться между
собой. Так, в растворе нуклеиновых кислот цепь с последовательностью GCTAT соединится с цепью CGATA. Особое внимание следует уделить тому, как
рестриктазы режут
297
ДНК. Вспомним, что большинство рестриктаз
разрезают симметричные палиндромные фрагменты и часто оставляют короткие
одинарные комплементарные концы цепей длиной в два-четыре основания. Например,
фермент SalI режет ДНК следующим образом:
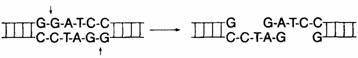
Все молекулы ДНК, порезанные этой
рестриктазой, будут иметь одинарные фрагменты с одинаковой последовательностью.
Эти концы могут соединяться посредством водородной связи, даже если они очень
короткие. Поэтому если порезать две ДНК разных организмов одним ферментом и
затем перемешать их, то можно получить много рекомбинантных молекул,
составленных из ДНК обоих видов.
На практике эксперимент, как правило,
сосредоточен на ДНК отдельного организма-донора,
например мыши или человека. Эту ДНК изолируют и проводят над ней различные
операции. Выделить ДНК из организма донора просто: достаточно разрушить клетки
и при помощи этанола выделить осадок ДНК. Затем донорскую ДНК обрабатывают
рестриктазой, чтобы она поделилась на мелкие фрагменты и эти мелкие фрагменты
вставляют в маленькие реплицирующие молекулы ДНК, служащие вектором (лат. vector - несущий); векторные молекулы переносят интересующую ученых ДНК, с
которой можно экспериментировать. Векторы могут реплицироваться, то есть
увеличивать количество донорских фрагментов и предоставлять много копий для
298
анализа. Наиболее часто использующиеся
векторы - плазмиды, с которыми мы
познакомились в гл. 10, то есть маленькие кольцевые ДНК, встречающиеся в
бактериях и часто переносящие гены устойчивости к антибиотикам. Плазмиды не
являются частью основного генома бактерий, поскольку встречаются штаммы как с
плазмидами, так и без них. Благодаря малому размеру и кольцевой структуре
плазмидных ДНК их легко отделить от геномной ДНК бактерий. В качестве векторов
также часто используют маленькие вирусные ДНК.
Порезав донорскую ДНК на фрагменты при
помощи рестриктазы, той же рестриктазой режут векторную молекулу, имеющую один
участок, соответствующий рестриктазе. Затем раствор двух ДНК смешивают и
получают некоторое количество рекомбинантных молекул ДНК - это и есть векторы
со встроенным донорским фрагментом:
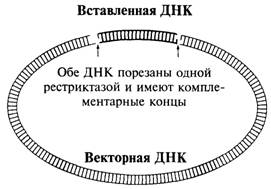
Такие вставки нестабильны, но если
добавить фермент ДНК-лигазу, то он соединит каркас молекул ДНК и сделает связь
прочной. В полученном растворе имеется большое количество векторных
299
302
Рис. 12.1. Для запуска
полимеразной цепной реакции отрезок ДНК нагревают до разделения его на две
цепи.
Затем к участкам на двух
цепях присоединяются праймеры, и цепи реплицируются при помощи ферментов,
устойчивых к высоким температурам. Получившиеся молекулы вновь разделяют
посредством нагревания, и цикл повторяется, в результате чего число молекул
удваивается
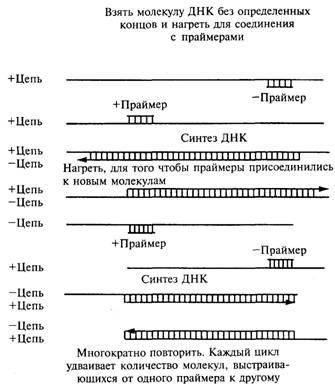
Некоторые доминантные дефектные аллели,
например тот, что вызывает болезнь Геттингтона, иногда проявляются на довольно
поздней стадии жизни. Метод PCR помогает
обнаружить их заранее.
303
Рис. 12.2. Метод ДНК-отпечатков
использован для установления невиновности одного из двух обвиняемых в
изнасиловании.
Образцы ДНК
подозреваемых А и В протестированы вместе с ДНК жертвы (ряд 6), образцом ДНК семени
с ее одежды (ряд 3) и образцом ДНК из влагалища (ряд 5).
Полученные результаты говорят о невиновности подозреваемого А и указывают на
то, что преступление мог совершить подозреваемый В

305
При помощи метода клонирования генов
можно создавать организмы с внедренными генами других организмов, и это,
пожалуй, самое захватывающее достижение современной генетики, используемое в
промышленных целях. Геном растения или животного можно изменять, добавляя
особые клонированные молекулы ДНК, благодаря которым организм приобретает новые
признаки.
Добавленный в геном ген называется трансгеном, а организм, полученный в
результате такой операции, называется трансгенным
организмом. В популярной литературе этот процесс известен под
названием генетическая модификация, но
это не совсем точное определение, так как полученные в результате традиционной
селекции организмы также в какой-то степени подвергаются генетической
модификации. Более точные термины 'генетически
модифицированные организмы' и 'генетически
модифицированные продукты' относятся исключительно к трансгенным
организмам.
Ключевое различие между трансгенными
организмами и организмами, полученными в результате селекции, состоит в том,
что трансгенная ДНК может быть перенесена практически из любого другого организма, и это невероятно
увеличивает возможности комбинирования признаков. При традиционной селекции
желательный аллель получали от особей того же или близкородственного вида.
Теперь же, если это необходимо в каких-то целях, гены рыб, например, можно
перенести в растение, а гены бактерий - в млекопитающее. Таким образом,
возможности преобразования ограничены теперь исключительно воображением, и
многих это заставляет задуматься, особенно если знать, насколько
306
309
вкладывают многомиллионные средства в
такие компании, как 'Сетус' (Беркли) и 'Байоджин' (Швейцария), обещающие
создать технологию, альтернативную микроэлектронике. Порой кажется, что
единственными ограничениями в этой области являются воображение и степень
талантливости исследователей. Перед недавними местными выборами в Онтарио
власти провинции заявили о многомиллионном проекте развития биотехнологий,
которые, судя по ожиданиям, должны обеспечить значительный прорыв в области
медицины, горного дела, лесного и сельского хозяйства, экологии и энергетики.
Помимо всего прочего, ожидается появление новых азотфиксирующих растений, а
также бактерий, которые очищали бы загрязнения, выделяли минералы и
вырабатывали алкоголь из промышленных отходов.
Среди разнообразных способов применения
трансгенных технологий особое место занимает генная
терапия. Если можно модифицировать растительные и животные
организмы, то что мешает применить те же методы для лечения наследственных
болезней? Технология рекомбинантных ДНК с самого начала подавала надежду на исправление
генетических нарушений посредством замены дефектного гена нормальным. Как
только был выделен ген, ответственный за развитие кистозного фиброза (CFTR), и его функция была подтверждена, начались
разработки средств лечения пациентов с кистозным фиброзом методами генной
терапии. Сначала нужно было определить, можно ли из отдель-
310
312
Все приведенные факты - примеры так
называемой соматической генной терапии, то
есть они применяются по отношению к телу (соме) пациента
в надежде, что получится достаточное количество клеток, способных выполнять
нормальные функции. Пациент может выздороветь, но риск передачи нежелательных
генов потомству все равно остается, потому что половые клетки таким образом не модифицируются.
Терапия половых клеток нацелена на
модификацию всего организма, включая и железы, вырабатывающие половые клетки.
Простейший (теоретически) способ состоит в том, чтобы модифицировать
оплодотворенную яйцеклетку, введя в нее подходящий трансген. Такого рода
процедура уже возможна и успешно проведена на опытных животных, например на
мышах. Но можно ли ее применить по отношению к человеку и, главное, стоит ли?
Это серьезный этический вопрос, и некоторые поборники нравственности
утверждают, что если соматическая генная терапия этична, то играть с
человеческим геномом и изменять генный набор наших потомков недопустимо,
поэтому подобные процедуры следует запретить.
Геномика - изучение всего генома
Последние достижения в области
секвенирования и развитие технических средств для обработки большого количества
клонов в библиотеке генов позволили ученым исследовать сразу весь геном
организма. Сейчас определены полные последовательности многих видов, в том
числе большинства так называемых модельных генетических организмов, таких как Е. coli, круглого червя Caenorhabditis elegans;
313
и, конечно, классического объекта
генетики, плодовой мушки Drosophila melanogaster. В 1990-х годах, несмотря на ряд неурядиц и разногласий, был начат
проект по исследованию человеческого генома ('Геном человека'), средства на
который выделил Национальный институт здоровья. В феврале 2001 года большая
группа исследователей во главе с Дж. Крэйгом Вентером из частной лаборатории
'Селера Дженомикс' сделали заявление о предварительной расшифровке
человеческого генома. Результат их работы был опубликован 16 февраля 2001 года
в журнале 'Science'.
Другая версия, которую представила группа
из Международного консорциума по секвенированию человеческого генома, была
напечатана 13 февраля 2001 года в журнале 'Nature'.
Временем зарождения геномики можно
считать середину ХХ века, когда генетики составили карты всех хромосом
модельных организмов, основываясь на частоте рекомбинаций (см. гл. 8). Однако
на этих картах были показаны лишь те гены, для которых были известны мутантные
аллели, и поэтому полными такие карты назвать нельзя. Полное секвенирование ДНК
позволяет выявить местонахождение всех генов организма, а также установить
последовательность оснований между ними.
Геномика делится на структурную и
функциональную. Структурная геномика ставит целью выяснить, где именно в
хромосомной ДНК расположены те или иные гены. Компьютерные программы распознают
типичные для генов начала и концы, отбирая те последовательности, которые,
вероятнее всего, и являются генами. Такие последовательности называют открытой рамкой считывания (open
314
Глава 13. ГЕНЕТИК В РОЛИ ДОКТОРА
ФРАНКЕНШТЕЙНА
В глазах современной общественности
генетики часто ассоциируются с образом героя романа Мэри Шелли 'Франкенштейн',
безумно увлеченного своей работой и создавшего ужасное чудовище. Генетиков
обвиняют в том, что они во что бы то ни стало, невзирая на финансовые траты,
стремятся приоткрыть завесу над тайнами жизни, создают вредные продукты и даже
нарушают мировой порядок. Того и гляди, жители всемирной деревни, уподобившись
своим собратьям из романа Шелли, вооружатся метафорическими вилами и пойдут на
штурм замка науки, топча по дороге поля генетически модифицированных продуктов
и выкрикивая лозунги протеста против 'новой генетики'.
Контроль над исследованиями
рекомбинантных ДНК
Споры о роли генетики начались задолго до
современного расцвета генной инженерии. Еще в 1970-х годах не только ученое
сообщество, но и широкая публика принялись обсуждать вопросы,
319
Генетически модифицированные организмы
Вопросы общественного влияния на генетику
и регулирования научных исследований в этой области, во многом не решены до сих
пор. По мере совершенствования микробиологических технологий
326
и методов появляются все новые и новые
поводы для беспокойства. В настоящее время речь уже идет не о том, чтобы
добавить несколько новых генов лабораторным бактериям. В наше время вполне
доступной стала технология создания в коммерческих целях генетически модифицированных организмов, то
есть трансгенных растений и животных с заданными признаками. Многие трансгенные
организмы до сих пор содержатся в лабораториях для тестирования, но некоторые
уже выпущены на рынок. Трансгенные технологии стали очередным поводом для
жарких споров, так как здесь сошлись интересы получения выгоды и сохранения
окружающей среды и здоровья людей. Вопрос сводится к тому, насколько безопасно
внедрять в организм чужую ДНК и насколько далеко можно предвидеть результаты
такого внедрения. Последние десять с лишним лет, в течение которых генетически
модифицированные продукты использовались достаточно широко, доказали, что эти
технологии не такие уж безвредные. Например, в некоторых случаях инсулин,
полученный от трансгенных бактерий, приводил к неблагоприятным последствиям.
Однако и особо опасных ситуаций еще не возникало. Национальный институт
здоровья пересмотрел некоторые свои рекомендации, и теперь генетики могут
пользоваться значительной свободой в своих экспериментах, хотя наиболее опасные
из них по-прежнему находятся под контролем. Вопросы по поводу модифицированных
организмов остаются, и стоит упомянуть хотя бы некоторые из них.
Первыми генетически модифицированными
организмами были бактерии центров кристаллизации льда и помидоры сорта Flavrsaver, выведенные
327
Одна из сторон возникшей проблемы -
научное просвещение. Как шутят агенты по продаже недвижимости, три ключевых
элемента, помогающих продать дом, - это его место, место и еще раз место. Точно
так же и в научном просвещении основные
329
три момента - это контекст, контекст и
еще раз контекст. Без знания контекста все новые открытия ученых подобны
воздушным шарам - красивым, но ничего не значащим для остального человечества.
Неприятие достижений современной генетики во многом коренится в том, что
неизвестен контекст их применения, и поэтому мы должны для начала рассмотреть
новые генетические технологии в свете технологии вообще.
Во всякой технологии есть свои
положительные и отрицательные стороны. Многие согласятся с тем, что
промышленная революция XIX века, основанная на научных достижениях в области
физики и химии, повысила уровень жизни в индустриально развитых странах. Однако
обратная сторона прогресса налицо. Например, промышленная революция подарила
человечеству двигатель внутреннего сгорания, с помощью которого передвигаются
автомобили и другие машины. Автомобиль люди любят за скорость, за то, что в нем
легко перемещаться в удаленные места, но при этом он наносит большой ущерб окружающей
среде и здоровью людей своими выхлопными газами, не говоря уже о том, что для
производства автомобилей добывают в шахтах металлы, выкачивают из недр земли
нефть; для строительства шоссе вырубают леса, уменьшая тем самым биологическое
разнообразие. С развитием транспортной сети города растут и еще более уменьшают
площадь нетронутых участков природы. Только в одной Канаде ежегодно около 16
тыс. смертей относят на счет загрязнения атмосферы транспортными средствами. К
этому количеству нужно добавить тысячи тех, кто гибнет в автомобильных авариях.
И эти смерти реальны, а не гипотетичны. Хотя
330
332
ствия применения и хлорфторуглеродов,
которые в первое время называли чудом современной химии. Эти вещества химически
инертны и служат прекрасными переносчиками химических веществ в аэрозольных
баллончиках. Никто не знал, что хлорфторуглероды будут накапливаться в верхних
слоях атмосферы и свободные радикалы хлора начнут разрушать озоновый слой.
Природа революционных технологий такова, что мы не можем предусмотреть всех
последствий их использования.
Аргументы против генетически
модифицированных продуктов
В ходе споров по поводу генетически
модифицированных продуктов был выдвинут ряд аргументов против их использования.
Мы перечислим здесь основные доводы противников, лежащие в основе их
рассуждений. Мы не призываем принять ту или иную сторону, а всего лишь
предлагаем внимательно подумать над этими аргументами, часть которых
заслуживает серьезного рассмотрения.
Сторонники трансгенных исследований
утверждают, что трансгенная ДНК - всего лишь ДНК, и генетики давно многое о ней
знают, поэтому, казалось бы, и нет повода для беспокойства. Однако, как ведет
себя ДНК, мы знаем на основании наблюдений и экспериментов, проводящихся внутри одного и того же вида. В ходе
трансгенных исследований ДНК одного вида переносят в клетки другого вида, и
было бы опасно рассуждать, что она бу-
333
344
Клонирование ДНК, как было сказано в гл. 12, представляет собой метод
генной инженерии, когда берут вектор и вставляют в него отдельный кусок ДНК,
после чего вектор проникает в клетки, производящие копии этой ДНК. Однако когда
биологи говорят о клонировании таких организмов, как овца или человек, они
подразумевают другой процесс: из оплодотворенной зиготы удаляется ядро, которое
заменяется ядром соматической клетки другого индивида. В удаленном ядре
содержались хромосомы от матери и отца, тогда как перенесенное ядро содержит
хромосомы одного индивида, донора, поэтому тот организм, который разовьется из
этой зиготы, должен быть генетически идентичен донору. Впервые метод
клонирования применили в исследованиях на лягушках и жабах для проверки
некоторых гипотез о развитии эмбриона. Формирование идентичных близнецов (двойняшек,
тройняшек и т. д.) представляет собой нечто вроде естественного клонирования
человека. Идентичные близнецы происходят от случайного разделения зиготы на две
клетки и более, которые затем развиваются в генетически идентичные организмы.
Овечка Долли развилась из ядра, взятого
из клетки молочной железы женской особи. (Свое имя Долли получила по имени
певицы в стиле кантри - пышногрудой Долли Партон.) Этой клетке позволили
делиться до той поры, пока она не подошла к такой стадии клеточного цикла,
когда ее ядро можно было ввести в зиготу.
Тот же метод успешно применили при
клонировании и других млекопитающих. В 2001 году некоторые лаборатории сообщили
об успешном клонировании людей, но получившиеся эмбрионы якобы
345
351
Геномика и здравоохранение
По мере развития геномики помимо проблемы генетически модифицированных продуктов возникают и иные вопросы, касающиеся здравоохранения. В настоящее время постоянно распознаются все новые и новые гены в ДНК человека, и становится возможным использовать маркеры, оповещающие о предрасположенности к тому или иному заболеванию. Вопрос заключается в том, до какой степени можно использовать эту генетическую информацию. В системах здравоохранения, построенных по образцу систем стран Европейского Сообщества, где каждый получает медицинскую помощь в соответствии со своими потребностями, подробное генетическое картирование может оказаться весьма полезным. Если знать, что человек расположен к тому или иному заболеванию, то можно провести ряд профилактических мер или облегчить протекание болезни, распознанной на ранней стадии. Но в других системах здравоохранения (вроде той, что принята в США), где каждый должен заключать частный страховой договор с компанией, основной целью которой является получение прибыли, существует опасность дискриминации людей по их геному. Эта проблема касается прежде всего не генетики, а общественно-политического использования генетической информации.
Глава 14. ИСТОЧНИК ПЕРЕМЕН: МУТАЦИИ
На протяжении всей истории человечества
рождение человека или животного с необычным внешним видом пробуждало страхи,
почтение или благоговейный ужас. В мифологиях многих культур говорится о
фантастических существах, созданных по прихоти богов или появившихся в силу
особого сочетания природных стихий. На самом деле это были люди и животные с
аномалиями развития или с искажениями наследственной информации; такие
организмы называются мутантами, а
искажение наследственной информации - мутацией. (Мутацией
называется также явление, приведшее к такому искажению.) Обычно под мутацией
подразумевают небольшие изменения в последовательностях нуклеотидных пар ДНК;
более крупные повреждения хромосом называются хромосомными
аберрациями. Поняв механизм мутаций, люди хотят научиться в какой-то
степени контролировать их, но в последнее время в окружающей среде под
воздействием человека появились новые факторы, вызывающие мутации, в первую
очередь радиоактивные и химические вещества, влияние которых мы начинаем
осознавать только сейчас.
В жутких фантастических фильмах и
рассказах обычно показаны вредные мутации, и складывает-
353
361
пространстве. Данные табл. 14.1
показывают количество фонового излучения, которое обычно получает человек за
год. Для сравнения приведены дозы излучений от искусственных источников - от
тера-
Таблица
14.1 Примерные дозы излучений
|
Источники излучения
|
Средняя доза на человека, мбэр
в год
|
|
|
Естественные источники
|
|
|
|
|
Космическое излучение
|
28
|
|
|
|
Земное излучение
|
26
|
|
|
|
Пища
|
28
|
|
|
|
Всего
|
82
|
|
|
Дополнительные источники
|
|
|
|
|
Рентгеноскопия
|
|
|
|
|
|
Пациенты
|
20
|
|
|
|
|
Персонал
|
< 0,15
|
|
|
|
Радиофармацевтические
препараты
|
|
|
|
|
|
Пациенты
|
2-4
|
|
|
|
Персонал
|
< 0,15
|
|
|
|
Потребительские товары
|
4-5
|
|
|
|
Служащие
|
|
|
|
|
|
Национальные лаборатории и
подрядчики
|
< 0,20
|
|
|
|
|
Промышленность
|
< 0,01
|
|
|
|
|
Военные
|
< 0,04
|
|
|
|
Выпадение осадков при испытании
ядерного оружия
|
4-5
|
|
|
|
Коммерческая атомная энергетика
|
|
|
|
|
|
Окружающая среда
|
< 1
|
|
|
|
|
Персонал
|
< 0,15
|
|
|
|
Разное (воздушный транспорт,
телевидение)
|
|
|
|
|
< 0,50
|
|
|
|
Всего
|
30-40
|
|
362
певтических и диагностических приборов до
атомных электростанций и люминесцентных циферблатов. За исключением медицинских
приборов для рентгеноскопии, искусственное излучение относительно невелико и
сравнимо с естественным.
Что представляют собой мутации?
Мутация - это изменение в ДНК. Некоторые
изменения происходят спонтанно, со временем. Например, молекулы ДНК теряют
пуриновые основания гуанин и аденин (депуринизация) с относительно высокой
скоростью. По примерным оценкам, клетка млекопитающего за сутки теряет около 10
тыс. пуриновых оснований. К счастью, в клетках имеются механизмы
восстановления, которые вставляют основания в нужные места или вырезают
искаженные последовательности и заменяют их вновь синтезируемыми.
Различные факторы также повреждают ДНК.
Некоторые повреждения удается восстановить при помощи специальных (репаративных)
ферментов, но другие повреждения становятся мутациями. В гл. 9 говорилось, что
мутаген профлавин вставляет или удаляет одно или несколько оснований и вызывает
так называемые мутации со сдвигом рамки, при которых механизм считывания
генетического кода сдвигается. Мутация может быть и заменой одного основания на
другое. Мутации такого рода образуются обычно при репликации ДНК, когда к
новому основанию прикрепляется не то основание, которое должно было
присоединиться изначально. Нормальные пары оснований, А-Т и G-C удер-
363
372
бомба с часовым механизмом, отсчитывающим
последние мгновения до ужасной генной катастрофы? К сожалению, пока никто не
может дать ответы на эти вопросы.
Хромосомы содержат гены, расположенные в
определенной последовательности. Фенотип организма зависит не только от тех или
иных генов, но и от того, как они расположены относительно других генов. На
первый взгляд это утверждение кажется странным. Если в ядре все хромосомы
перемешаны, то, казалось бы, местоположение гена роли не играет; главное - то,
что он присутствует. Поэтому генетики поначалу немало удивились, узнав, что
перестройки хромосом могут привести к серьезным изменениям в экспрессии генов.
Почему это происходит, во многом остается загадкой. О регуляции генов,
упомянутой в гл. 11, мы знаем еще не очень много, но нам известно, что
положительная или отрицательная регуляция генов осуществляется участками ДНК,
расположенными от них на некотором удалении. Поэтому изменение местоположения
гена может включить его в иную регуляторную систему. В любом случае
перестановки последовательностей ДНК подобно их потерям и вставкам воздействуют
на развитие половых клеток и на рост организма.
Хромосомные аберрации происходят
вследствие физических разрывов хромосом и неправильного воссоединения их
фрагментов. Делеция, или утрата
участка хромосомы, происходит вследствие двух разрывов или отрыва конечного
участка. Если ото-
373
375
хромосом; многие нарушения настолько
незаметны, что их нелегко распознать при современном уровне диагностики. По
приблизительным оценкам, сегодня более чем у 10% детей имеются нарушения,
которые требуют основательного лечения или хирургического вмешательства еще в
младенчестве или в раннем детском возрасте. В это число не входят прекращения
беременности на самой ранней стадии развития эмбриона, когда некоторые женщины
даже не догадываются, что были беременны. Так что хромосомные аберрации отнюдь
не редки, и они служат серьезным источником беспокойства и страданий.
Под электронным микроскопом хромосомы
человека выглядят как свитые в многочисленные петли куски толстой веревки.
Каждая хромосома представляет собой длинную, непрерывную цепь ДНК, в скрученном
виде окруженную особыми белками и молекулами РНК. Поначалу исследования
хромосом (цитогенетика) ограничивались изучением хромосом
растений и насекомых с малым числом крупных хромосом. У млекопитающих обычно
бывает наоборот - большое количество небольших хромосом. С 1920-х до середины
1950-х годов было распространено мнение, что у человека 48 хромосом. (Когда Д.
Сузуки, один из авторов этой книги, учился в колледже, ему говорили, что у
кавказцев 48 хромосом, а мужчины - представители желтой расы имеют хромосомный
набор Х0 и 47 хромосом!). Но в 1956 году шведские ученые Трийо и Леван провели
ряд ис-
376
следований и обнаружили, что в самых
хорошо подготовленных клетках при самых благоприятных условиях всегда
наблюдается 46 хромосом. С тех пор общепризнанным стало число 46. У приматов
количество хромосом сравнимо с количеством хромосом у человека, у макак-резусов
их 42; у шимпанзе, горилл и орангутанов - 48.
В гл. 5 мы уже говорили, что для
получения кариотипа раствор белых кровяных клеток обрабатывают колхицином. На
четких фотографиях видны различные хромосомы, которые можно распределить по
порядку. Всем хромосомам человека присвоены порядковые номера, начиная с самой
длинной (рис. 14.2). После длины самая заметная характеристика - положение
центромеры, то есть участка, в котором хроматиды сцеплены между собой и к
которому присоединяются нити веретена. Хромосомы с центромерами, расположенными
приблизительно посередине, называются метацентрическими,
как, например, первые три хромосомы человека. У акроцентрических хромосом центромеры
расположены ближе к одному из концов и делят их на две неравных части, или
'плечи' (как, например, хромосомы с 16 по 18). Бывают также телоцентрические хромосомы с центромерами
практически на конце, у человека таких нет. У некоторых хромосом имеются сателлиты, небольшие участки, присоединенные к
основной хромосоме такой тонкой нитью, что ее почти не видно.
Более отчетливое изображение можно
получить посредством окрашивания. При окрашивании раствором Гимза становятся
видны кольцевые полосы, или Гимза-диски; под
действием других красителей появляются флуоресцентные полосы. Такие процедуры
помогают опознать хромосомы среди
377
Глава 15. ЭВОЛЮЦИОННАЯ ГЕНЕТИКА
Генетик Феодосий Добржанский однажды
сказал: 'Все в биологии имеет смысл только в свете эволюции'. Все биологи,
объективно взирающие на окружающий мир, соглашаются с тем, что огромное
разнообразие видов на нашей планете появилось в результате естественных
процессов, которые мы называем эволюцией. Почему столько людей убеждены в
истинности эволюционной теории? Некоторые циники и религиозные критики
сравнивают их убеждение с верой в Бога и утверждают, что это своего рода
научная религия. Однако вера в религиозном понимании и вера в научную теорию
отличаются друг от друга, и главное их отличие заключается в том, как устроена
наука.
Наука не имеет дело с истиной. Она имеет
дело с гипотезами, правильность которых можно
оспорить. Ученый начинает с того, что замечает ряд необъяснимых
явлений и старается найти им рациональное толкование. Обычно научное
рассуждение строится по образцу, который философ Н. Р. Хэнсон назвал ретродукцией: 'Вот необъяснимое явление; но
это явление станет объяснимым, если X
верно; следовательно, я заявляю, что X верно'.
Формулировка подходящего объяснения требует некоторого скач-
387
ка воображения, и по этой причине в науке
есть доля творчества, как и в любом другом виде человеческой деятельности.
Однако при этом наука накладывает ряд ограничений на то, что можно считать X. Объяснение X
должно принадлежать к сфере обычной
физической реальности. Ученый не может ссылаться на сверхъестественные силы,
такие как Бог, демон или какого-либо рода магия. Самое главное ограничение
состоит в том, что гипотеза должна быть проверена эмпирически,
то есть в ходе опытов и экспериментов, она должна предсказывать
наблюдаемые последствия. После формулировки гипотезы ученые приступают к
эмпирической проверке предсказываемых ею последствий.
Далекие от науки люди часто полагают, что
посредством опытов и экспериментов ученые стараются найти подтверждения своей
гипотезы. На самом деле они стараются доказать, что она ложна. Конечно, в
глубине души ученые надеются, что они правы, но хитрость логики состоит в том,
что этого они доказать никогда не смогут. Допустим, имеется некоторая гипотеза
Г, которая предсказывает последствия П; суть ее выражена формулой 'Если Г, то
П'. Мы проводим опыты, чтобы удостовериться в наличии П. Предположим, что мы
наблюдаем П. Можно ли на основании этого построить следующее умозаключение:
'Если Г, то П; П верно, значит, верно и Г'? Нет, нельзя. Такая схема умозаключения
ошибочна, ведь нельзя по следствию судить о причине. (Попробуйте рассуждать
следующим образом: 'Если солнце состоит из горящего навоза, то оно должно быть
горячим. Солнце горячее, следовательно, оно состоит из горящего навоза'.) Но
предположим, что мы не наблюдаем П. Тогда мы
388
с полной уверенностью можем утверждать: 'Если Г, то П; П не верно, значит, Г
не верно'. Как говорил философ Карл Поппер, гипотеза должна быть фальсифицируемой, то есть она должна
предсказывать такие последствия, на основании проверки которых ее можно опровергнуть. Если результаты эксперимента не
согласуются с гипотезой, то ее либо отвергают, либо подвергают пересмотру. Если
результаты согласуются с гипотезой, то она на некоторое время становится
рабочей. Из этого вовсе не следует, что она верна, просто на данный момент она
предлагает самое подходящее объяснение фактам. По мере выдвижения все новых и
новых гипотез мы совершенствуем их и составляем общую картину, то есть
разрабатываем теорию - более широкую систему идей, объясняющих реальность.
Конечно, и теории не являются истиной в последней инстанции, они лишь
предлагают более правдоподобные объяснения большого количества явлений. К
такого рода теориям относится и теория эволюции. Она выдержала многочисленные
испытания и доказала свою надежность. Но мы не должны забывать, что теория
эволюции рассматривает два вопроса: 1) происходит
ли эволюция (правда ли то, что современные виды возникли таким
образом?) и 2) как она происходит?
Сомневаться в том, что эволюция происходит, было бы отрицанием почти всей
современной биологии, как правильно подметил Добржанский. За последние 150 лет
накоплены горы свидетельств и экспериментальных подтверждений, которые
подкрепляют (точнее, которым не удается опровергнуть) идею происхождения одних
видов от других.
389
Другой вопрос эволюционной теории - о
механизме эволюции - не решен до сих пор, и о нем идут многочисленные споры в
современной науке. Правда, почти никто не сомневается в том, что эволюция
зависит от естественного отбора, как
объясняется далее. Авторство этой идеи приписывается Чарльзу Дарвину и Альфреду
Уоллесу, хотя некоторые ученые и до них высказывали подобные мысли. Оба
естествоиспытателя много путешествовали, наблюдали за разнообразием природы
нашей планеты, исследовали животных, описывали экзотические растения и
окаменелости. В потрясающем многообразии им удалось разглядеть сходство и общие
законы. Дарвин проиллюстрировал общие принципы на примере передней конечности
позвоночных, имеющей у всех одинаковое строение - длинную кость в верхней
части, две в нижней, несколько костей в запястье и пять пальцев, состоящих из
нескольких коротких костей:
Что может быть более любопытным, нежели
то, что рука человека, предназначенная для хватания, передняя конечность крота,
предназначенная для рытья, нога лошади, плавник дельфина и крыло летучей мыши
устроены по одному образцу и состоят из схожих костей, расположенных в похожем
порядке?
Такое сходство органов, выполняющих
разные функции, но имеющих общую схему строения, называется гомологией. И до Дарвина анатомы подмечали
сходство в строении органов разных животных, но объясняли их с точки зрения
традиционной религии, хотя некоторые предполагали нечто вроде эволюции. Дарвин
и Уоллес объяснили гомологию тем, что разные животные происходят от одного общего
предка. Они предположили, что пер-
390
воначальная группа организмов разделилась
на несколько популяций, оказавшихся в разных средах обитания. В каждой
популяции, вероятнее всего, выживали и давали потомство организмы с
наследуемыми признаками, позволявшими им легче приспособиться к среде обитания.
Так происходил естественный отбор приспособленных
организмов. За множество поколений случайные изменения в геноме привели к
появлению новых признаков, а естественный отбор закрепил наиболее полезные из
них. Так произошла прогрессивная дивергенция (расхождение) видов и появились
современные формы, такие как человек, летучая мышь и дельфин.
Доказательства того, что разные организмы
действительно произошли от общего предка посредством постепенного изменения,
поступают из разных источников. Пожалуй, одно из самых сильных доказательств -
гомология на всех уровнях биологической организации. Исследования разных групп
организмов предоставили множество примеров гомологии видимых структур, таких
как конечности или череп. Причем благодаря окаменелостям гомологичное строение
органов можно проследить не только у современных, но и у вымерших животных. Эти
окаменелости можно опознать как предков некоторых современных видов, как
переходное звено от одного вида к другому, подтвердив тем самым родство
современных форм жизни. Собрав вместе всю информацию о той или иной группе
животных, можно построить так называемое филогенети-
391
В широком плане эволюция охватывает три
процесса: макроэволюцию, специализацию и микроэволюцию. Макроэволюция подразумевает совокупность всех
процессов, благодаря которым в прошлом существовали различные типы организмов,
и им на смену приходили другие организмы. О масштабе макроэволюции можно
судить, в частности, по окаменелостям. Специализация означает развитие двух
видов и более с различными признаками на месте одного общего предка. Микроэволюция подразумевает небольшие
изменения внутри одного вида. Отсюда логично следует вопрос: а что же такое,
собственно, вид? На этот вопрос нет простого ответа. Понятно, что словом 'вид'
обозначают некую отдельную разновидность организмов, но достаточно заглянуть в
любую книгу о деревьях, цветах, птицах, насекомых и других организмах, чтобы
увидеть поразительное сходство многих видов между собой. Где провести границу?
В случае с организмами, имеющими пол, наиболее очевидное (и весьма спорное)
определение вида гласит, что это группа организмов, способных давать общее
потомство. С представителями другого вида эти организмы скрещиваться и давать
потомство не могут. Иногда представители одного и того же вида сильно
отличаются друг от друга по внешнему виду, но потомство
394
396
ный вид может развиваться на протяжении
долгого времени и приобретать новые признаки. Это называется филетической эволюцией. Но основной процесс
эволюции - вымирание: эволюция - это
не история о том, как появлялись и выживали новые виды, а печальное
повествование, как вымирали старые. Виды существуют на протяжении определенного
времени, от нескольких сотен тысяч до нескольких миллионов лет, а после
вымирают, так как не могут приспособиться к изменившимся условиям окружающей
среды (при этом к факторам окружающей среды относятся и другие виды, которые
также эволюционируют).
Делить аллели генов на дикие и мутантные,
как мы это делали, знакомясь с основами генетики, не совсем правильно, и такое
деление может привести к неправильному представлению об эволюции. Исследования
природных популяций показывают, что не у всех членов популяции общий генотип,
который мы условно называем диким. На самом деле, во многих популяциях
наблюдается значительное генетическое разнообразие. Добржанский с коллегами
провели исследования диких дрозофил на юго-западе США и обнаружили, что среди
них бывают носители нескольких инверсионных вариантов
каждой из хромосом. (Инверсия - это поворот одного из участков хромосомы.) В
слюнных железах плодовых мушек бывают гигантские хромосомы с четким рисунком
черных и белых полос, которые видны под микроскопом. Таким образом легко
сравни-
397
400
ляции наследственного заболевания,
вызываемого одним аллелем. Возьмем для примера такое аутосомное рецессивное
заболевание, как фенилкетонурия, которое в популяции встречается с частотой q2. Если в определенной популяции от фенилкетонурии страдает один человек
на 10 тыс., то q2 = = 1/10 000. Отсюда следует, что q должно
быть равно квадратному корню из 1/10 000, то есть 1/100. Так как р + q = 1, то р = 99/100.
Тогда согласно формуле Харди-Вайнберга частота гетерозиготных носителей 2pq = 2 х 99/100 х 1/100
= 1/50 (приблизительно). Эти подсчеты показывают, что
гетерозиготные носители встречаются гораздо чаще (приблизительно один на 50
человек), чем гомозиготные больные. Знание частоты гетерозигот очень помогает
при генетическом консультировании. Зная данные о распространении гетерозигот,
можно также постараться устранить методом отбора рецессивный аллель из
популяции, как будет описано далее.
Наиболее противоречивый вывод из теории
эволюции Дарвина заключался в предположении, что человек произошел от обезьяны.
Представители христианской религии приняли эту идею с неодобрением, потому что
она противоречит библейской истории о том, что все живые существа на Земле
созданы Богом за короткий промежуток времени. Поначалу не существовало особых
доказательств, что человек действительно произошел от обезьяны, за исключением
некоторого сходства частей тела и органов. Антропологи начали искать
'недостающее зве-
401
но', то есть окаменелые останки существ,
которые можно было бы назвать промежуточной формой между обезьянами и
человеком. За последующие 150 лет было найдено несколько 'промежуточных' видов,
и антропологи получили достаточно сведений, чтобы проследить эволюцию человека
и его предков, которых назвали гоминидами.
Самые древние останки были найдены в
Африке, и сейчас с большой уверенностью можно утверждать, что гоминиды изначально
обитали на этом континенте. О самых ранних гоминидах, называемых Australopitecus afarensis, почти ничего не известно, за исключением того, что они жили около 4-3,7
млн лет назад. Они были довольно невысокими (около 1,2 м ростом), ходили на
двух ногах и имели небольшой мозг. Их сменили два других вида австралопитеков -
Australopitecus africanus и
Australopitecus robustus, которые были крупнее и обладали мозгом большего размера. Последние
австралопитеки, жившие около 2,5-1,5 млн лет назад, по всей видимости,
соседствовали с первыми видами, принадлежавшими к роду Homo (человек) - Homo habilis, то есть с человеком умелым, названным так потому, что он умел
изготовлять примитивные орудия труда. Человек умелый был выше своих предков (1,5
м), и у него был сравнительно большой мозг (около половины объема мозга
современного человека). Затем появился более высокий Homo erectus (человек прямоходящий) с более развитым мозгом,
который жил около 1-0,25 млн лет назад. Почти 250 тыс. лет назад появился
современный вид человека - Homo sapiens (человек разумный). Однако среди этого вида на ранней
стадии было несколько разных форм, например не-
402
Миграция и разнообразие Homo sapiens
Благодаря секвенированию ДНК людей по
всему миру удалось построить филогенетическое дерево человечества. Корни этого
дерева, как свидетельствуют окаменелые останки, уходят в Африку. Большинство
биологов придерживаются теории, согласно которой Homo sapiens из Африки несколькими волнами мигрировал по всему
свету. Дивергенция среди разных групп во время миграций и после них привела к
появлению так называемых человеческих рас, если слово 'раса' определять как
'группу людей, первоначально обитавшую в определенном регионе и обладающую
внешними признаками, заметно отличающими ее от другой группы людей'.
Поскольку история контактов между
различными группами людей доказывает, что все они могут скрещиваться между
собой, все мы принадлежим к одному и тому же виду. Иногда в это не так уж легко
поверить, особенно если, например, сравнить темнокожих низкорослых пигмеев из
Центральной Африки и высоких европейцев со светлой кожей. В свое время
антропологи пытались на основе видимых различий построить классификацию
человечес-
403
Как уже говорилось в гл. 1, мысль об
улучшении человеческого рода зародилась давно, по крайней мере, в
древнегреческом обществе классического периода. Но особое внимание она
привлекла в последние десятилетия XIX и в начале ХХ века, когда
408
начала распространяться теория об
естественном отборе и развития классической генетики. Первым идею о проведении
специальных мер по улучшению человечества высказал двоюродный брат Чарльза
Дарвина, Фрэнсис Гальтон, который и ввел в научный обиход термин 'евгеника',
что означало 'хорошее рождение (порода)'. Исходя из того, что человек выводит
породы животных и сорта растений с нужными ему признаками, Гальтон высказал
предположение, что для улучшения признаков людей также нужен искусственный
отбор. Эту идею подхватили многие ученые и общественные деятели того времени, в
том числе и писатель Джордж Бернард Шоу. В Европе и Северной Америке был
осуществлен ряд евгенических программ. При этом одни касались наследственных
болезней и дефектов, а другие были связаны с представлением о превосходстве
одних рас над другими.
К тому времени выяснилось, что многие
заболевания и дефекты связаны с одним аллелем гена и передаются по классическим
законам Менделя. Но сторонники евгеники утверждали, что по наследству
передаются и многие другие синдромы, такие, как слабоумие, бродяжничество и
криминальность. Утверждалось, что для процветания общества нужно искоренить из
популяции эти ненужные признаки. В некоторых европейских странах, а также в США
и Канаде, были созданы евгенические комиссии, оценивающие необходимость
евгенических мер в том или ином конкретном случае. В результате были
стерилизованы десятки тысяч людей, часто без особых причин. В некоторых странах,
например Канаде и Швеции, подобные мероприятия проводились до конца 1970-х
годов. В 1995 году в Китайской Народ -
409
1
Mark Schorer, William Blake: The Politics
of Vision (New York; Vintage, 1959).
2
Joseph Haberer, 'Politicalization in Science', Science, 178, 1977. P. 713-723.
3 Theodore Roszak, The Monster and the Titan: Science, Knowledge, and
Gnosis, Daedalus, Summer 1974, pp. 17-34.
1
Robert Graves, 'Introduction', in New
Larousse Encyclopedia of Mythology (London: Hamlyn, 1959, p. vii).
2 Гомер, Илиада. Песнь V, стихи 265-274. Перевод Н. Гнедича.
3 Вергилий. Георгики. III. Пер. С. Шервинского.
4
Stuart Queen and Robert Habenstein, The Family in Various Cultures.(Philadelphia: J. B. Lippincott, 1974), pp. 158-9.
1 Единица атомного веса иногда
называется дальтоном в честь
английского химика Джона Дальтона.
1 Термином признак обозначается отличительная особенность
организма, передающаяся по наследству, например цвет глаз и волос или окраска
цветка растения. Под чертой мы здесь
подразумеваем отдельную форму признака, например лиловые цветки растений.
414
1 Заметьте, что и клетки, и хромосомы
размножаются делением; сначала они удваиваются, затем две половины расходятся.
В биологии принято условно называть образованные в результате деления клетки
дочерними, а хромосомы - сестринскими.
2 Kennedy McWhirter, 'XYY Chromosome
and Criminal Acts', Science, 164,
1969, p. 1117.
3
Georgetown Law Journal, 'Note:
The XYY Chromosome Defense', 57, 1969, pp. 892-922.
4 Barbara J. Culliton, 'Parents'
Rights: Harvard Is Site of Battle over X and Y Chromosomes', Science, 186, 1974, pp. 715-17.
5 H. Bentley Glass, 'Endless Horizons
or Golden Age', Science, 171, 1971, pp.
28ff.
6
Ernest B. Hook, 'Behavioral Implications of the Human XYY Genotype', Science, 179, 1973, pp. 139-50.
1 Гаррод понимал значимость сделанного им
открытия, но, как это часто бывало в истории науки, другие ученые его не поняли,
и труды Гаррода вновь открыли лишь несколько десятилетий спустя, когда генетика
продвинулась достаточно далеко.
2
Robert Guthrie, 'Mass Screening for Genetic Disease', in Medical Genetics Today, ed. D. Bergsma (Baltimore: John Hopkins Press, 1974).
1 Некоторые штаммы Е. coli, особенно 0157:Н7, в последнее время
печально прославились тем, что вызывают серьезные (порой даже летальные)
желудочно-кишечные заболевания, но это редкие исключения. В лабораториях
исследуются безвредные штаммы Е. coli.
2 Felix d'Herelle, 'The
bacteriophage', Science News, 14,
1949, pp. 44ff.
1 Некоторые фаги, растущие на
К, являются rII+-ревертантами, испытавшими обратную мутацию,
но частота реверсии
415
измеряется
независимо и вычитается из общего числа; она гораздо меньше частоты
рекомбинации.
' Laurie Garrett, The Coming Plague (New York: Farrar, Strauss &
Giroux, 1994).
2
Bernard Dixon, 'Antibiotics and Advertisers', New Scientist, 10 April 1975, p. 58.
3 Theodore Friedmann and Richard Roblin, 'Gene Therapy for Human
Diseases?', Science, 175, 1972, pp. 949-55.
4
Stanley Rodgers, 'Gene Therapy for Genetic Disease?', Science, 178, 1972, pp. 648-9.
Глава тринадцатая
1 'Монсано' была одной из первых компаний,
начавших активно выступать против Рэйчел Гарсон, опубликовавшей книгу
'Безмолвный источник' (Silent Spring). Эта
книга пробудила интерес широкой публики к пестицидам, загрязняющим окружающую
среду, и способствовала развитию движения за охрану окружающей среды.
2 Здесь не место обсуждать
экономические теории, но тем, кто доверяет принципам традиционной экономики,
будет интересно узнать, что в последнее время развивается школа так называемой
экологической экономики, которая отвергает многие традиционные идеи и старается
рассматривать экономическую деятельность в более широком биологическом
контексте.
Глава четырнадцатая
1 Доза излучения измеряется в
зависимости от его ионизирующего воздействия на сухой воздух; единицей служит рентген. Бэр - это так называемый биологический эквивалент рентгена, то есть
доза излучения, которая вызывает то же количество повреждений, что и один
рентген рентгеновского и гамма-излучений. Миллибэр равен одной тысячной бэра.
Авторадиография - метод получения снимка радиоактивных материалов посредством их
воздействия на фотографический раствор; там, где раствор проявляется,
образуется темное пятно.
Агар -
полисахарид, выделяемый из морских водорослей и используемый в качестве плотной
питательной среды.
Аденин -
одно из пуриновых оснований ДНК или РНК.
Активный центр -
участок или углубление в молекуле фермента, где происходит химическая реакция.
Алкаптонурия -
нарушение метаболизма, характеризуемое умственной отсталостью вследствие
накопления в организме гомогентизиновой кислоты (алкаптона), которая темнеет на
воздухе.
Аллель -
одна из нескольких форм гена.
Аллостерический белок - белок с двумя активными центрами, каждый из которых
связывается со специфическим лигандом. В зависимости от того, с каким лигандом
он связан, меняются структура и активность.
Амбер-мутация - мутация, в результате которой образуется нонсенс-кодон (стоп-кодон) (UAG), приводящий к преждевременной остановке синтеза
белка.
Амин -
органическое соединение, имеющее в своем составе аминогруппу NH2.
Аминный конец -
конец пептидной цепи со свободной аминной групой.
Аминоацильная тРНК - транспортная РНК, соединенная с аминокислотой, которую она переносит
для синтеза белка.
Аминогруппа -
химическое соединение NH2.
417
Аминокислота -
мономер белка, органическое соединение, имеющее в своем составе аминогруппу NH2
и карбоксильную группу COOH.
Анафаза -
фаза митоза и мейоза, при которой хромосомы или хроматиды расходятся по разным
полюсам.
Анеуплоид -
организм, имеющий избыток или недостаток хромосом по сравнению с нормальным (эуплоидным)
хромосомным набором.
Антиген -
чужеродное тело, попавшее в организм (обычно птицы или млекопитающего), в ответ
на внедрение которого вырабатываются антитела.
Антикодон -
последовательность молекулы транспортной РНК, дополнительная к
последовательности кодонов молекулы матричной РНК.
Антитело -
молекула, которую вырабатывают определенные клетки (обычно птиц и млекопитающих)
в ответ на антигены; обычно способна связываться с антителом и нейтрализовывать
его.
Ауксотроф -
мутантный организм, неспособный синтезировать одно или несколько жизненно
необходимых веществ.
Аутосома -
обычная хромосома, в отличие от половой хромосомы (X или Y).
Бактериофаг -
вирус бактерий.
Белок -
полимер, состоящий из аминокислот.
Биоинформатика -
наука, анализирующая структуру и функцию генома, особенно методами
компьютерного моделирования и программирования.
Биосинтез -
часть метаболизма, ответственная за синтез молекул, из которых состоит
организм.
Бластула -
стадия развития эмбриона, полое шаровое скопление клеток.
Бляшка -
небольшое чистое пятно посреди слоя клеток, выросших в чашке с питательной
средой; причиной появления этого пятна служат вирусы.
Быстрообменивающаяся метка - метка, добавляемая к исследуемым клеткам или
организмам на короткий
418
промежуток времени с целью изучения процессов в этих
клетках.
Вакуоль -
небольшая, ограниченная мембраной клеточная структура в виде пузырька,
переносящая материалы из одного места в другое внутри клетки.
Валентность -
способность атома образовывать определенное число связей с другими атомами.
Вектор -
вирус, плазмида или другая частица, с помощью которой осуществляется перенос
ДНК.
Веретено деления - структура клетки, состоящая из микротрубочек, которая во время митоза
или мейоза растаскивает хромосомы по разным полюсам.
Видообразование - процесс образования новых биологических видов.
Вирион -
полостью сформированная вирусная частица, существующая вне клетки и состоящая
преимущественно из нуклеиновой кислоты (генома) и белковой оболочки (капсида).
Вирулентный фаг - бактериофаг, способный размножаться только в ходе литического цикла.
Вирус -
биологическая частица, отличающаяся от организма и способная размножаться
только внутри функционирующей клетки; состоит из генома и на определенной
стадии развития образует внеклеточные вирионы; см. вирион.
Водородная связь - слабая связь, удерживающая вместе молекулы; при этом один
положительно заряженный ион водорода связывает два относительно отрицательно
заряженных атома, например: -О...Н-О- или- N...H-О-, здесь эта
связь обозначена точками.
Врожденное нарушение метаболизма - генетическое нарушение, приводящее к неспособности
синтезировать фермент, который участвует в метаболической реакции.
Вырожденный код - код, в котором одной единице информации соответствует несколько
символов. Генетический код является вырожденным, потому что не-
419
которым аминокислотам соответствует более одного
кодона.
Газон - слой
бактерий, выращенный на питательной среде, в котором можно наблюдать бляшки
фагов и вирусов; в более общем смысле слой любой клеточной культуры.
Гаплоид (гаплоидный) - организм или клетка, имеющие одинарный набор
хромосом; см. диплоид.
Генетически модифицированный организм (ГМО) - трансгенный организм, в который перенесли один или
несколько генов неродственного ему организма.
Генная терапия половых клеток - предполагаемое направление генной терапии, при
котором дефектные аллели будут заменяться в половых (репродуктивных) клетках,
что предотвратит передачу их потомству.
Геном -
совокупность всех генов и регуляторных сигналов, определяющих структуру и
активность вирусов или организмов; генетическая информация, закодированная в
нуклеиновой кислоте.
Геномная библиотека - набор фрагментов ДНК одного вида организмов, порезанных эндонуклеазой
и перенесенных в векторы.
Генотип -
набор аллелей определенных генов в конкретном организме; ср. фенотип.
Гетерозигота (гетерозиготный) - диплоидный организм, имеющий два разных аллеля
одного гена; см. гомозигота.
Гидроксильная группа - химическое соединение ОН.
Гидрофильный -
буквально 'любящий воду'; гидрофильным называется химическое вещество,
растворимое в воде; см. гидрофобный.
Гидрофобный -
буквально 'боящийся воды'; гидрофобным называется химическое вещество,
неспособное растворяться в воде; см. гидрофильный.
Гликоген -
полисахарид, образованный полимеризированными молекулами глюкозы; основной
запасной углевод животных и человека.
420
Гомеотический ген - регуляторный ген, мутация которого приводит к значительным изменениям
в развитии.
Гомозигота (гомозиготный) - диплоидный организм, имеющий два одинаковых аллеля
одного гена; ср. гетерозигота.
Гомологичные хромосомы - в диплоидном организме - хромосомы, имеющие
одинаковую форму и по преимуществу одинаковый набор генов.
Гомология -
сходство в строении (анатомическом или молекулярном) между двумя биологическими
видами.
Гомункулус -
в истории науки - гипотетическое миниатюрное существо, содержащееся в каждом
сперматозоиде или яйцеклетке, из которого за время беременности развивается
ребенок.
Градиент плотности - раствор (часто хлорида цезия, CsCl) в пробирке, помещаемой в центрифугу, в котором
наблюдается постепенное увеличение плотности в направлении центробежной силы
центрифуги. Молекулы в таком растворе распределяются по своей плотности или
размеру.
Гуанин -
одно из пуриновых оснований ДНК или РНК.
Дезоксирибоза -
вариант сахара рибозы с пятью атомами углерода, в котором атом С2 вместо атома водорода удерживает гидроксильную
группу ОН.
Делеция -
хромосомная мутация, при которой теряется фрагмент хромосомы.
Денатурация -
изменение структуры молекулы, например белка, при котором она теряет свою
биологическую активность.
Детерминация -
процесс эмбрионального развития, при котором определяется судьба клеток; см. дифференциация.
'Дикий тип' -
для лабораторных организмов - стандартный фенотип или аллель, отличающийся от
мутантного фенотипа и мутантных аллелей.
421
Дипептид -
молекула, состоящая из двух аминокислот, соединенных пептидной связью.
Диплоид (диплоидный) - организм или клетка, имеющие двойной набор хромосом.
См. гаплоид.
Дифракция рентгеновских лучей - рассеяние рентгеновских лучей кристаллическими
объектами, на основе которого возможно определить структуру этих объектов.
Дифференциация -
процесс эмбрионального развития, при котором клетки принимают специфическую
форму и начинают выполнять определенную функцию; см. детерминация.
ДНК-лигаза -
фермент, образующий фосфоэфирную связь между двумя фрагментами нуклеиновой
кислоты, то есть выстраивающий из них цепь.
ДНК-пластинка -
небольшая стеклянная пластинка с нанесенными на нее одноцепочечными молекулами
ДНК, представляющими собой фрагмент генома. Используется для выявления
комплементарных молекул РНК.
ДНК-полимераза -
фермент, реплицирующий ДНК и соединяющий нуклеотиды в цепь ДНК.
Доминантный -
один из нескольких аллелей гена называется доминантным, если он проявляется в
гетерозиготном состоянии; см. рецессивный.
Донор - 1)
при спаривании бактерий - клетка (Hfr или F+),
передающая ДНК реципиенту (F~); 2) в генной
инженерии - организм, из которого извлекается фрагмент ДНК для последующего
изучения или переноса в другой организм.
Дупликация -
хромосомная мутация, при которой фрагмент хромосомы удваивается.
Естественный отбор - процесс выживания и воспроизведения организмов, наиболее
приспособленных к условиям среды. Благодаря естественному отбору виды занимают
определенную экологическую нишу.
Зародышевая клетка - клетка, из которой образуется гамета.
422
Зигота - клетка, образовавшаяся в результате слияния
сперматозоида и яйцеклетки.
Зонд -
молекула, обычно небольшая нуклеиновая кислота, которая служит меткой для
нахождения отдельного гена или генетического элемента.
Инверсия -
хромосомная мутация, при которой фрагмент хромосомы отрывается и вставляется в
обратном порядке.
Индукция - 1)
экспрессия гена посредством инактивации белка-репрессора; 2) при эмбриональном
развитии - процесс, при котором один тип клеток вызывает изменения в клетках
другого типа.
Интерфаза -
фаза цикла эукариотической клетки, во время которой не происходит митоза.
Интрон -
некодирующая последовательность оснований внутри одного гена; см. экзон.
Информационная РНК - см. матричная РНК.
Информация -
мера упорядоченности или совокупность сведений о чем-либо.
Ион - атом
или молекула с положительным или отрицательным зарядом.
Капсид -
защитная оболочка вириона (вирусной частицы).
Карбоксильная группа - химическое соединение COOH, называемое еще кислотной
группой, потому что атом водорода стремится отделиться в виде иона Н+.
Карбоксильный конец - конец полипептидной цепи со свободной карбоксильной группой.
Кариотип -
схема хромосомного набора организма, получаемая в результате специальной
обработки клеток, фотографирования и распределения хромосом по порядку.
Карта сцеплений - схема хромосомы, на которой показаны участки генов и других
генетических элементов, определяемых в зависимости от степени их сцепления.
Картирование методом делеции - процесс составления генетической карты при помощи
мутантов по деле-
423
ции. Если две и более мутации пересекаются, то они не
могут рекомбинировать.
Катализатор -
вещество, убыстряющее химическую реакцию, но не преобразующееся в результате
этой реакции.
Килобаза -
тысяча оснований, нуклеотидов или нуклеотидных пар.
Клетка -
фундаментальная биологическая единица, представляющая собой сложный комплекс
биологических молекул и ионов, растворенных в воде, окруженных плазматической
мембраной.
Клеточный цикл -
ряд процессов, включающий в себя репликацию ДНК и заканчивающийся делением
клетки.
Клон - 1)
группа организмов, происшедших от общего предка путем бесполого размножения (например,
в результате клеточного деления); 2) организм, созданный в результате переноса
ядра клетки одного организма в яйцеклетку без ядра другого организма; 3)
участок ДНК, внедренный в вектор и многократно реплицированный (клон ДНК).
Клон ДНК -
вектор (и клетка, в которой он реплицируется), содержащий фрагмент внедренной
ДНК.
Клонирование -
эксперимент по созданию клона (см. клон 2 и 3).
Ковалентная связь - химическая связь, образованная двумя и более атомами, имеющими одну
или несколько общих пар электронов. Для разрыва такой связи требуется
значительное количество энергии.
Кодирующая цепь - цепь ДНК, последовательность оснований которой идентична
последовательности мРНК, за исключением того, что место тимина занимает урацил.
Кодоминантный -
два аллеля кодоминантны, если в гетерозиготном организме они выражаются в
равной степени.
Кодон -
единица генетического кода, которая состоит из трех оснований и кодирует синтез
одной отдельной аминокислоты.
424
Колинеарность -
тип прямого соответствия между последовательностями нуклеиновой кислоты и
аминокислот в белке, синтез которого она кодирует.
Колония -
группа клеток, таких, как бактерии, выращиваемых в виде клона из одной
клетки-предшественника, обычно на поверхности чашки с агаром.
Колхицин -
химическое вещество, останавливающее митоз в метафазе; позволяет составить
кариотип.
Комплементарная ДНК - молекула ДНК, созданная с молекулы РНК при помощи фермента обратной
транскриптазы (ревертазы).
Комплементарные молекулы - молекулы, строение которых позволяет им связываться
друг с другом. В частности, комплементарными могут быть основания нуклеиновых
кислот; см. комплементарные основания.
Комплементарные основания - нуклеотидные основания, образующие стабильные пары
друг с другом: аденин с тимином или урацилом и гуанин с цитозином.
Комплементарные цепи - цепи ДНК, последовательности оснований которых
комплементарны друг другу; см. комплементарные
основания.
Конъюгация -
процесс соединения двух клеток (особенно бактерий), при котором происходит
обмен их ДНК.
Крахмал -
полисахарид, состоящий из двух остатков глюкозы, соединенных между собой связью
альфа 1:4; накапливается в растениях в качестве запасного питательного
вещества.
Кроссинговер -
процесс, при котором две молекулы нуклеиновой кислоты (в хромосомах)
разрываются и обмениваются фрагментами.
Лиганд -
молекула, связанная с активным центром белка.
Лизис -
процесс, при котором инфицированная фагом (или, в общем смысле, вирусом) клетка
разрушается и освобождает фаг, находящийся внутри нее.
Лизогения -
состояние, при котором бактерии содержат в себе профаг - неинфекционную форму
фага.
425
Лизогенные бактерии - бактерии, которые находятся в состоянии лизогении.
Липиды -
класс органических веществ, к которым принадлежат жиры; обычно их молекулы
представляют собой длинные цепи углеводородов.
Литический цикл - процесс, при котором бактериофаг размножается в клетке хозяина и
разрушает эту клетку.
Локус -
местоположение гена или регуляторного элемента на хромосоме или на генетической
карте.
Макроэволюция -
смена одних видов другими, наблюдаемая на примере ископаемых остатков.
Маркер -
нуклеиновая кислота с мутацией или другой заметной особенностью, используемая
для определения положения гена или других генетических элементов.
Матрица -
структура или молекула, которая служит образцом для синтеза комплементарной
структуры или молекулы.
Матричная РНК (мРНК) - молекула РНК, последовательность кодонов которой
соответствует последовательности аминокислот в том или ином белке.
Матричная цепь -
одна из двух цепей ДНК, которая служит матрицей для синтеза РНК и,
следовательно, последовательность оснований которой комплементарна
последовательности РНК.
Мезодерма -
у эмбриона животных - первичная ткань, из которой образуется большинство
внутренних органов; см. эндодерма, эктодерма.
Мейоз -
процесс, при котором из диплоидной клетки образуются клетки с гаплоидным
набором хромосом (половые клетки).
Мембрана -
клеточная структура, состоящая из белков и липидов, которая служит границей
клетки. Внутри нее находятся другие клеточные структуры, такие, как ядро и
митохондрии.
Метаболизм -
общая совокупность химических процессов, происходящих в организме.
426
Метаболит -
промежуточный продукт метаболического пути.
Метаболический путь - серия химических реакций, осуществляемых с помощью ферментов,
благодаря которым одни вещества преобразуются в другие.
Метафаза -
во время митоза или мейоза - стадия, на которой хромосомы выстраиваются посреди
делящейся клетки.
Метка -
радиоактивный атом, а также некоторые другие атомы и молекулы, которые
внедряются в некоторые сложные вещества; с помощью меток легко следить за
процессами и определять структуру клеток или молекул.
Метод 'отпечатков пальцев' (фингерпринтинг) - определение индивидуальной структуры фрагментов ДНК
при помощи особого фермента и разделения этих фрагментов в геле.
Микроэволюция -
небольшие изменения, происходящие внутри отдельного биологического вида.
Митоз - в
эукариотических клетках - процесс, при котором ядро делится на два ядра с
идентичным набором хромосом. Как правило, сопровождается делением клетки.
Митохондрии -
небольшие вытянутые тельца в эукариотических клетках, в которых питательные
вещества преобразуются в энергию и создается запас энергии в пригодной для
потребления клеткой форме.
Модель - в
науке - схема или описание какого-либо процесса, его условное изображение.
Молекулярная масса - масса молекулы, выраженная в атомных единицах массы. За единицу
принята масса атома водорода (точнее, одна двенадцатая массы атома углерода).
Мономер -
небольшая молекула, одна из составных частей полимера.
Моносахарид -
отдельная молекула сахара, мономер полисахаридов.
427
Мутаген -
вещество или явление, такое, как излучение, которое повышает частоту мутаций.
Мутация - 1)
процесс, в результате которого происходят изменения в геноме; 2) собственно
изменение генома.
Направление репликации - направление, в котором происходит синтез ДНК (от
3'-конца к 5'-концу).
Направление транскрипции - направление, в котором происходит синтез РНК с одной
из цепей ДНК.
Нонсенс-кодон (стоп-кодон) - см. терминирующий
кодон.
Нонсенс-мутант -
мутант, у которого место кодирующего кодона занимает нонсен-кодон, или
стоп-кодон (UGA, UAG или UAA), сигнализирующий об окончании синтеза полипептида и
прерывающий синтез раньше времени.
Нуклеотид -
мономер нуклеиновой кислоты, состоящий из сахара (рибозы или дезоксирибозы),
связанный с азотистым основанием и фосфатом.
Оогенез -
процесс образования яйцеклетки.
Ооцит -
клетка, из которой в процессе мейоза образуется яйцеклетка.
Оператор -
регуляторный элемент, примыкающий к гену или блоку генов; имеет участок, к
которому присоединяется регуляторный белок.
Оперон -
один или несколько генов, экспрессия которых регулируется оператором.
Органелла -
отдельная структура клетки, выполняющая специфическую функцию.
Органические соединения - химические вещества, молекулы которых состоят из
одного или нескольких атомов углерода, а также водорода, кислорода и/или азота;
органические вещества входят в состав организмов.
Основание -
1) вещество, реагирующее с ионами водорода. Например, основанием является
гидроксид натрия NaOH, который в воде разлагается
на ион Na+
и
428
ион ОН"; ион ОН" соединяется с ионом
водорода Н+ и образует очередную
молекулу воды; 2) в более узком смысле азотистый компонент нуклеиновой кислоты.
Открытая рамка считывания - предполагаемый ген, распознаваемый как серия
кодонов между типичным начальным кодоном и типичным терминирующим кодоном,
достаточно большой для кодирования полипептида.
Пангенезис -
теория (ныне устарелая), согласно которой сперматозоиды и яйцеклетки вбирают в
себя вещества со всех частей и органов тела; предполагалось, что таким образом гаметы
передают информацию о признаках родительского организма потомству.
Пептидная связь - связь С-О-N-Н, посредством которой
соединяются две аминокислоты, образуя пептид, или белок.
Первичная структура - последовательность аминокислот в цепи белка.
Пермиссивное условие - условие, при котором летальный мутант получает
возможность расти, например низкая температура для мутантов, чувствительных к
теплу; см. рестриктивное условие.
Пиримидиновые основания - азотистые основания нуклеиновых кислот - цитозин,
тимин и урацил, представляющие собой кольцо из шести атомов; см. пуриновые основания.
Питательная среда - смесь питательных веществ, в которой выращивают некоторые организмы.
Плазмиды -
молекулы ДНК, находящиеся в бактериях и некоторых других организмах, способные
к самостоятельному размножению; передают отдельные признаки и иногда могут
переносить свою копию в другую клетку, хотя, как правило, существуют отдельно
от генома клетки.
Плейотропный аллель - аллель, обусловливающий проявление более одного фенотипического
признака.
429
Повторяющаяся ДНК
- ДНК, содержащая повторяющиеся короткие последовательности и образующая
большую долю хромосом эукариот.
Подвид - отдельная группа внутри биологического вида,
отличающаяся особыми признаками и ареалом распространения.
Полимер -
молекула, состоящая из многих идентичных или схожих небольших молекул (мономеров).
Полимераза -
фермент, осуществляющий синтез полимера.
Полимеразная цепная реакция - процесс увеличения количества молекул ДНК
посредством многократной репликации.
Полиморфизм длины рестрикционных фрагментов - различие между ДНК разных индивидов, обнаруживаемое
в результате обработки ДНК определенным рестрикционным ферментом (рестриктазой).
Полинуклеотид - нуклеиновая кислота; молекула,
состоящая из множества нуклеотидов.
Полипептид - белок; молекула, состоящая из многих
связанных между собой аминокислот.
Полирибосома -
структура, состоящая из молекулы мРНК, присоединившейся к нескольким рибосомам.
Полисахарид -
молекула, состоящая из множества связанных между собой сахаров (моносахаридов).
Полуконсервативная репликация - модель репликации ДНК, при которой цепи двойной
молекулы разделяются и вдоль каждой изначальной цепи выстраивается новая.
Полюса деления -
при мейозе или митозе противоположные стороны делящейся клетки, к которым
расходятся хромосомы и в которых образуются новые ядра.
Полярное тельце - один из крохотных нефункциональных продуктов, образующихся при
оогенезе (мейозе ооцита).
Посев -
распределение микроорганизмов и вирусов в чашке Петри в целях их выращивания и
изучения.
430
Преформация -
теория, согласно которой в сперматозоиде или яйцеклетке существует крохотный,
но уже сформировавшийся организм.
Промотор -
участок молекулы ДНК (хромосомы), к которому присоединяется молекула
РНК-полимеразы и с которого она начинает транскрипцию.
Прототроф -
организм, такой, как бактерии или грибы, способный синтезировать необходимые
для своей жизнедеятельности вещества (аминокислоты, нуклеотиды и др.) из
простых углеводородных соединений, таких, как сахара.
Профаг - разновидность
умеренного фага, такого, как лямбда (λ) или P1, находящегося в лизогенном
состоянии; обычно кольцевая молекула нуклеиновой кислоты, существующая отдельно
или встроенная в геном бактерии.
Профаза -
первая фаза митоза или мейоза, во время которой происходит уплотнение хромосом
и разрушение ядерной оболочки.
Профлавин -
краситель, молекулы которого связываются с молекулой ДНК и вызывают мутации
вставки или делеции.
Псевдовирион -
вирусная частица, содержащая клеточную ДНК вместо вирусной ДНК, посредством
которой осуществляется осуществлять трансдукция. Пунктирной линией обозначена
водородная связь.
Пуриновые основания - азотистые основания нуклеиновых кислот - аденин или гуанин, в которых
кольцо из шести атомов связано с кольцом из пяти атомов; см. пиримидиновые основания.
Рамка считывания - воображаемая рамка длиной в три основания, по которой происходит
считывание кодирующих триплетов вдоль нуклеиновой кислоты.
Распознавание -
процесс, благодаря которому органическая молекула соединяется с другой молекулой
или с определенной последовательностью оснований нуклеиновой кислоты.
431
Расщепление признаков - процесс, при котором аллели гена переходят в разные
гаметы.
Ревертант -
мутантный организм, в котором произошла очередная мутация, вернувшая его к 'дикому
типу'.
Регуляторный ген - ген, кодирующий производство регуляторного белка.
Рекомбинант -
организм, имеющий набор аллелей, отличающийся от набора каждого из родителей.
Рекомбинантная ДНК - ДНК, полученная в результате обработки исходной ДНК определенной
рестриктазой и вставки ее фрагмента в векторную ДНК, порезанную той же
рестриктазой.
Реплика -
копия молекулы нуклеиновой кислоты, идентичная первоначальной.
Репликация -
процесс, при котором из одной молекулы нуклеиновой кислоты образуется две
идентичные молекулы.
Репрессор -
регуляторный белок, регулирующий экспрессию одного или нескольких генов
посредством связывания с определенным оператором.
Рестриктаза (эндонуклеаза рестрикции) - фермент, который разрезает ДНК на фрагменты в местах
определенных последовательностей.
Рестриктивное условие - условие, при котором летальный мутант не может
расти, например высокая температура для мутантов, чувствительных к теплу; см. пермиссивное условие.
Рестрикционная карта - карта хромосом, сделанная на основе сравнения
фрагментов, порезанных различными рестриктазами.
Рецептор -
белок, имеющий центр связывания с определенным лигандом, функция этого белка
состоит в распознавании этого лиганда, благодаря чему осуществляется
необходимая реакция.
Рецессивный аллель - один из нескольких аллелей гена называется рецессивным, если он
проявляется только в гомозиготном состоянии и не проявляется в гетерозиготном;
см. доминантный аллель.
432
Реципиент -
при конъюгации бактерий - клетки F-, спаривающиеся с клетками Hfr или F+.
Решетка Пеннета - таблица, с помощью которой определяют возможные результаты
генетического скрещивания. По вертикали отмечают гаметы одного родителя, а по
горизонтали - другого. Получившиеся в ячейках таблицы сочетания указывают на
возможный генотип потомства.
Рибоза -
сахар с пятью атомами углерода, входящий в состав нуклеотидов.
Рибосома -
клеточная структура, представляющая собой комплекс белков и молекул РНК, в
которых происходит синтез белков.
Рибосомная РНК (рРНК) - молекула РНК, входящая в состав рибосом.
РНК-полимераза -
фермент, с помощью которого с молекулы ДНК синтезируется молекула РНК; фермент,
осуществляющий транскрипцию.
Родословная -
схема родственных связей между организмами, на примере которых можно изучать
законы наследственности и распределения признаков среди потомства.
Серповидноклеточная анемия - наследственное заболевание, при котором красные
кровяные клетки приобретают серповидную форму и закупоривают небольшие кровяные
сосуды.
Синтения -
особенность, при которой у разных организмов имеются схожие или идентичные гены
на одних и тех же участках хромосом.
Соматическая генная терапия - процесс исправления генетических нарушений
посредством внедрения нормальных генов в ткань, поврежденную вследствие
дефектных генов.
Соматическая клетка
- в многоклеточном организме - любая клетка (например, клетка печени, мышцы или
кожи), за исключением половых (гамет или клеток, которые подвергаются мейозу).
433
Сперматиды -
клетки, образуемые в ходе мейоза, из которых впоследствии получаются
сперматозоиды.
Сперматогенез -
процесс образования сперматозоидов.
Сперматозоид -
зрелая мужская гамета.
Сперматоцит -
клетка, которая в ходе мейоза делится на несколько гамет (сперматозоидов).
Спирт -
органическое соединение, имеющее в своем составе гидроксильную группу ОН.
Стволовая клетка - клетка многоклеточного организма, которая сохраняет способность
развиваться в клетки различного типа.
Структурная геномика - наука, изучающая структуру генома.
Субстрат -
химическое вещество, подвергающееся действию фермента и преобразуемое в
результате реакции.
Супрессор -
вторичная мутация, которая восстанавливает фенотипические признаки, утраченные
в результате первой мутации.
Сцепленные гены - гены, находящиеся на одной хромосоме и наследуемые, как правило,
вместе.
Телофаза -
последняя стадия митоза или мейоза, во время которой происходит образование
новых ядер вокруг отдельных наборов хромосом.
Терминирующий кодон - нонсенс-кодон (стоп-кодон) (UAG, UGA или UAA), который служит сигналом для прекращения синтеза
полипептидной цепи.
Тестовое скрещивание - скрещивание, предпринимаемое в целях определения
генотипа индивида доминантного фенотипа, который скрещивается с гомозиготным
индивидом рецессивного фенотипа. Если первый индивид гомозиготный, то все
потомство будет с доминантным фенотипом, но если он гетерозиготный, то часть
потомства будет обладать рецессивным фенотипом.
Тимин - одно
из пиримидиновых оснований ДНК.
Ткань - в
многоклеточном организме - структура, состоящая из множества однотипных клеток,
выполня-
434
ющих определенную функцию (например, мышцы, кожа,
нервная ткань).
Тотипотентная клетка - клетка, способная развиться в любой из
специализированных видов клеток; см. стволовая
клетка.
Трансген -
ген, перенесенный в геном неродственного организма.
Трансгенный организм - организм, геном которого был изменен путем внедрения
чужеродного гена.
Трансдукция -
процесс, при котором вирус (как правило, бактериофаг) переносит клеточную (геномную)
ДНК из одной клетки в другую.
Транскрипт -
молекула РНК, синтезируемая в ходе транскрипции ДНК; последовательность
оснований транскрипта комплементарна последовательности оснований ДНК.
Транскрипция -
процесс синтеза матричной РНК, переносящей информацию отдельного гена.
Транслокация -
хромосомная аберрация, при которой часть одной хромосомы присоединяется к
другой, не гомологичной ей хромосоме.
Трансляция -
процесс синтеза белка на основе информации РНК; см. транскрипция.
Транспозон -
генетический элемент, способный перемещаться в геноме с одного места на другое.
Транспортная РНК (тРНК) - небольшая молекула РНК, переносящая аминокислоту для
синтеза белка.
Трансформация -
процесс преобразования клетки, обычно бактерии, после внедрения в нее ДНК.
Ультрацентрифуга - аппарат для разделения молекул смеси или взвеси под действием
центробежной силы.
Умеренный бактериофаг - бактериофаг, способный существовать в лизогенном
виде.
Урацил -
одно из пиримидиновых оснований РНК.
Уридин -
рибонуклеотид, содержащий урацил.
Фаг - вирус
бактерий.
435
Фенилкетонурия -
генетически обусловленное нарушение, характеризуемое умственной отсталостью
вследствие неспособности организма перерабатывать финилалалин.
Фенотип -
совокупность признаков и свойств организма; см. генотип.
Фенотип, зависимый от условий, - фенотип, который не позволяет организму выживать в
одних (неблагоприятных) условиях и позволяет нормально функционировать в других
(разрешающих) условиях.
Фиксация азота -
химический процесс, при котором атмосферный азот N2
превращается в форму, в которой его могут потреблять растения, в виде аммиака NH3
или нитрата NO3-.
Филетическая эволюция - эволюционный процесс, в ходе которого вид постепенно
меняет свои признаки.
Филогенетическое дерево - схема предполагаемых эволюционных связей между
разными видами.
Фингерпринтинг -
см. метод 'отпечатков пальцев'.
Фосфат -
химическая комбинация РO4, встречаемая в нуклеиновых кислотах и других
соединениях.
Хиазма -
точка переплетения гомологичных хроматид во время мейоза, в которой они могут
разрываться; в результате такого разрыва хроматиды могут обмениваться своими
частями.
Химическая реакция - процесс, при котором атомы и молекулы взаимодействуют друг с другом и
образуют другие молекулы.
Химическая связь - взаимодействие между двумя атомами, благодаря которому они образуют
соединение (см. ковалентная связь).
Хлоропласт -
органелла фотосинтезирующей эукариотической клетки (например, растения), в
которой происходит фотосинтез.
Хромомера -
небольшое утолщение на хромосоме.
Хромосома -
структура, состоящая из ДНК в комплексе с гистонами и другими белками, а также
молекулами
436
РНК; один из самых основных клеточных компонентов,
содержащий генетическую информацию. Хромосомные аберрации - изменения
структуры или количества хромосом.
Целлюлоза -
полимер глюкозы, важная составляющая тканей растений (особенно древесины).
Центриоль -
структура в клетках животных (и некоторых других), располагающаяся на полюсах
делящейся клетки и направляющая движение хромосом к полюсам.
Центромера -
участок хромосомы, в котором хроматиды соединяются друг с другом.
Цистрон -
функциональная единица, соответствующая гену, но выделяемая на основании
комплементационного теста.
Цитозин -
одно из пиримидиновых оснований ДНК или РНК.
Частота аллелей - в популяционной генетике соотношение нескольких аллелей одного гена (или
типа хромосом).
Частота мутаций - мера вероятности того, что произойдет мутация; в общем случае
вероятность того, что отдельный ген подвергнется мутации за определенное
количество делений клетки.
Чувствительный к температуре - мутант, не способный нормально функционировать при
высокой температуре.
Чувствительный к холоду мутант - мутант, нормально функционирующий при средних
температурах, но ненормально функционирующий при низких температурах.
Штамм -
специфический тип микроорганизмов или вирусов, обычно имеющий определенный
генотип и выращиваемый в исследовательских целях.
Эктодерма -
у эмбриона животных первичная ткань, из которой образуются наружные ткани (кожа)
и основная часть нервной системы; см. эндодерма,
мезодерма.
437
Ядерная оболочка - комплекс из двух мембран, образующий границу ядра в эукариотической
клетке.
Ядро - в
клетках эукариот органелла, в которой содержатся хромосомы.
Яйцеклетка -
женская половая клетка, из которой в результате оплодотворения может развиться
новый организм. Содержит гаплоидный набор хромосом.
Hfr-штамм -
штамм Е. coli со встроенным в геном
фактором F, вследствие чего
клетки этого штамма служат донорами при конъюгации.
Neurospora - обычная хлебная плесень красного цвета, используемая для генетических
экспериментов.
R-плазмида -
плазмида, содержащая гены, определяющие устойчивость к одному или нескольким
антибиотикам.
S-период -
период, в течение которого реплицируется ДНК хромосом эукариотической клетки.
Х-хромосома -
у млекопитающих и некоторых других животных хромосома, определяющая пол
индивида; у самок имеет парную Х-хромосому; у самцов парной хромосомой служит Y-хромосома.
Y-хромосома -
у млекопитающих и некоторых других животных определяет пол индивида (мужской);
составляет пару с Х-хромосомой.
ПРЕДИСЛОВИЕ.............................................................5
Глава первая. ГЕНЕТИКА:
ПРОШЛОЕ,
НАСТОЯЩЕЕ И
БУДУЩЕЕ...................................9
Поиски порядка и
смысла...................................... 12
Современный образ
науки...................................... 16
Перспективы современной
генетики.....................24
Глава вторая. ОТ
МИФА К СОВРЕМЕННОЙ
НАУКЕ.....................................................................28
Примитивный интерес к наследственности.........31
Одомашнивание растений и животных
в зеркале мифа.........................................................32
Научные теории
наследственности........................39
Откуда берутся
дети?...............................................44
Глава третья. ЧТО
ЖЕ ИМЕННО ПЕРЕДАЕТСЯ
ПО НАСЛЕДСТВУ?................................................52
Строение
клеток......................................................54
Молекулярная
структура.........................................59
Рост и
биосинтез.....................................................70
Ферменты.................................................................73
Синтез
полимеров...................................................78
Клетки как фабрики по самовоспроизводству
и
самообновлению...................................................80
Глава четвертая. РЕВОЛЮЦИОННОЕ
ОТКРЫТИЕ: ЗАКОНЫ МЕНДЕЛЯ......................81
Открытия
Менделя..................................................83
Родословные............................................................88
Другой пример: тестеры и нетестеры.....................95
Группы
крови...........................................................98
Множественные аллели и доминантность............103
Тестовые
скрещивания...........................................104
439
Вероятность.............................................................105
Два гена и
более.....................................................108
Первый закон Менделя и определение
отцовства.................................................................112
Ответы на вопросы о группах крови.....................114
Глава пятая. ХРОМОСОМЫ,
РАЗМНОЖЕНИЕ
И
ПОЛ.....................................................................115
Клетки и
размножение...........................................115
Митоз и клеточный
цикл.......................................118
Кариотип.................................................................121
Мейоз......................................................................124
Мейоз и законы
Менделя......................................130
Местонахождение генов.........................................132
Половые
хромосомы..............................................133
Нерасхождение
хромосом......................................136
Мужчины XYY: 'хромосома
преступности'.........139
Попытки определения пола...................................143
Глава шестая. ФУНКЦИЯ
ГЕНОВ.............................148
Гены и нарушения
метаболизма...........................148
Гены и
ферменты...................................................151
Белки и информация.............................................153
Исправление наследственных нарушений............161
Глава седьмая. НАСЛЕДСТВЕННЫЙ
МАТЕРИАЛ,
ДНК.................................................169
Бактерии..................................................................170
Первые шаги...........................................................174
Вирусы.....................................................................175
Бактериофаги..........................................................178
Эксперимент Херши-Чейз...................................181
Строение
ДНК........................................................184
Модель ДНК и
генетика........................................189
Проверка
модели....................................................191
Глава восьмая. СТРОЕНИЕ
ГЕНА..............................196
Распределение
генов..............................................196
440
Кроссинговер внутри
генов...................................205
Генетика
фагов.......................................................206
Тонкая структура
гена............................................208
Комплементация и определение границ гена......209
Что же такое
ген?...................................................212
Рестрикционные ферменты и палиндромы..........214
Секвенирование ДНК............................................216
Рестрикционное
картирование..............................219
Глава девятая. РАСШИФРОВКА
КОДА ЖИЗНИ ...224
Как строятся
белки?...............................................228
Молекулы РНК: инструменты
для синтеза
белка....................................................229
РНК-транскрипция................................................233
Трансляция..............................................................237
Сложные гены эукариот........................................239
Генетический
словарь............................................243
Колинеарность генов и
белков..............................246
Терминирующие
кодоны.......................................248
Универсальность кода............................................249
Глава десятая. НАСЛЕДСТВЕННОСТЬ
В МИРЕ
БАКТЕРИЙ.............................................251
Бактерии-мутанты..................................................251
Пол у Е. coli............................................................253
Плазмиды................................................................257
Факторы резистентности и устойчивость
к
антибиотикам.......................................................259
Лизогения................................................................265
Гены, переносимые
вирусом.................................267
Трансдукция и геном
человека..............................268
Отбор
мутантов.......................................................272
Глава одиннадцатая. РЕГУЛЯЦИЯ ГЕНОВ
И РАЗВИТИЕ
ОРГАНИЗМА................................274
Регуляция генов у
бактерий...................................276
Белки, которые
связываются................................280
Регуляция генов
эукариот......................................282
Эмбриональное развитие в общих чертах.............284
441
Регуляция по времени и развитие крыла цыпленка.................................................................288
Регуляция по положению и развитие тела мухи ....290
Формирование глаза мухи..........292
Глава двенадцатая. ВМЕШАТЕЛЬСТВО В СТРОЕНИЕ ДНК: ВОЗВРАЩЕНИЕ
ЭПИМЕТЕЯ?..........................................................296
Рекомбинантная ДНК и рестриктазы...................297
Изучение отдельных клонированных фрагментов..............................................................300
Трансгенные
организмы........................................305
Генная
терапия.......................................................310
Геномика - изучение всего
генома......................313
Глава тринадцатая. ГЕНЕТИК В РОЛИ ДОКТОРА
ФРАНКЕНШТЕЙНА.............................................319
Контроль над исследованиями рекомбинантных
ДНК............................................319
Генетически модифицированные организмы......326
Технологии в контексте.........................................329
Аргументы против генетически модифицированных
продуктов..............................333
Этические аспекты
клонирования........................344
Ответственность
ученых.........................................348
Геномика и здравоохранение................................352
Глава четырнадцатая. ИСТОЧНИК ПЕРЕМЕН:
МУТАЦИИ.............................................................353
Частота
мутаций.....................................................355
Мутации у людей....................................................357
Излучение...............................................................358
Что представляют собой
мутации?........................363
Система восстановления
ДНК..............................367
Генетические последствия
радиации....................369
Хромосомные
аберрации.......................................373
Хромосомы
человека..............................................376
Анеуплоидия...........................................................379
Дупликация и
делеция...........................................380
442
Инверсии............................................................. 382
Транслокации................................................... 384
Глава пятнадцатая. ЭВОЛЮЦИОННАЯ
ГЕНЕТИКА.............................................................387
Доказательства
эволюции......................................391
Эволюция как
процесс...........................................394
Популяционная
генетика.......................................397
Эволюция
человека................................................401
Миграция и разнообразие Homo sapiens................403
Евгеника..................................................................408
ПРИМЕЧАНИЯ...........................................................414
СЛОВАРЬ
417
'Грандиозный мир'
Формат 84x108/32, переплет

Томпсон М.
Философия науки / Пер. с англ. -
2003. - 304 с.
Книга представляет собой краткий обзор
вопросов, которые изучает философия науки. Основное внимание автор уделил
истории научной мысли, важнейшим научным и философским концепциям, а также
проблемам, связанным с влиянием науки на жизнь общества.

Мур П. Астрономия с Патриком Муром / Пер.
с англ. - 2004. - 368 с: ил.
Книга известного популяризатора
астрономии Патрика Мура будет интересна всем. Его талант говорить просто о
сложном подвигнет новичка к овладению навыками наблюдения за звездным небом, а
профессионала заставит по-новому взглянуть на предмет и научит излагать его
доступным, понятным языком. Таким образом, у вас в руках равно и пособие для
начинающих, и методическое руководство для лекторов и преподавателей.

Томпсон М.
Восточная философия / Пер. с англ. - 2002. - 384 с: ил.
Доктор Мел Томпсон, специалист по
философии, вероисповеданиям и этике, посвятил свою книгу описанию
фундаментальных религиозных и философских традиций. Он знакомит с индуизмом,
джайнизмом и буддизмом, которыми одарила человечество индийская философия,
конфуцианством и даосизмом, сформировавшими китайскую школу мысли,
религиозно-философскими традициями тантризма и дзэн-буддизма.

Великие
мыслители о великих вопросах: Современная западная философия / Пер. с англ. -
2001. - 400 с.
В этой книге внимание двадцати двух известных западных
ученых сосредоточено на пятнадцати вечных вопросах. Философы размышляют о
происхождении Вселенной, существовании Бога, души вне тела, о вероятности жизни
после смерти. Здесь вы встретите широкий спектр взглядов и сможете проследить
связь между современной мыслью и представлениями об истине, очевидными для большинства
людей.
'Грандиозный мир'
Формат 84x108/32, обложка

Брейтот Дж. 101
ключевая идея: Физика. - Пер.
с англ. - 2001.
- 256 с.
Эта книга знакомит читателя с физикой,
привлекая внимание к знакомым предметам, раскрывая их незнакомые стороны. Здесь
объясняется 101 ключевая идея великой науки, расширяющей наши знания о мире.

Дженкинс М. 101
ключевая идея: Эволюция. -
Пер. с англ. -
2001. - 240 с.
Читатель познакомится с эволюцией,
описываемой в доступной форме. В книге объясняется 101 ключевой термин, часто
встречающийся в литературе по данной отрасли знаний.

Митчелл П.
101 ключевая идея: Экология. - Пер. с англ. -
2001. - 224 с.
Цель этой книги - доступным и
увлекательным образом познакомить читателя с экологией. Здесь объясняется 101
ключевой термин, часто встречающийся в литературе по данной отрасли знаний.

Оливер П. 101
ключевая идея: Философия. - Пер.
с англ. - 2001.
- 240 с.
Эта книга знакомит читателя с философией.
Здесь объясняется 101 ключевой термин, часто встречающийся в литературе по
данной отрасли знаний.
предлагает вниманию читателей
Словари-справочники
Формат 60x90/16, переплет

Словарь мифов
/ Пер. с англ. - 2001. - 432 с: ил.
Словарь мифов, подготовленный британскими
специалистами, представляет собой собрание мифотворчества народов мира, в
котором ярко и красочно представлены персонажи славянской, кельтской, японской,
китайской, индуистской, шумеро-аккадской, греческой, римской, ацтекской и
многих других мифологий. Новый взгляд на мифологических героев, многие из
которых хорошо нам знакомы, будет интересен всем, кто неравнодушен к
загадочному миру человеческих фантазий и верований.
Кордуэлл М.
Психология. А - Я: Словарь-справочник / Пер. с англ. - 2003. - 448 с: ил.
Психолог и преподаватель Майк Кордуэлл
создал свой словарь-справочник по психологии, максимально приблизив к нуждам
учащихся. Каждый термин или понятие сопровождается кратким определением, так
что читатель может воспользоваться им при подготовке ответа на учебное или
экзаменационное задание.


Лоусон Т.,
Гэррод Д. Социология. А - Я: Словарь-справочник / Пер. с англ. - 2000. - 608
с: ил.
В справочник включены все основные
термины на русском и английском языках, с которыми приходится сталкиваться в
процессе изучения дисциплины, а также изложены базовые принципы и концепции
социологической науки. Содержание статей проиллюстрировано примерами, рисунками
и графиками.

Уолл Н.,
Маркузе Я., Лайнз Д., Мартин Б. Экономика и бизнес. А - Я: Словарь-справочник /
Пер. с англ. - 1999. - 624 с: ил.
Словарь-справочник, созданный
преподавателями английских университетов, представляет собой пособие с
алфавитным порядком расположения статей, максимально облегчающим его
практическое применение. Каждый термин или понятие сопровождается кратким
определением, так что читатель может воспользоваться им при подготовке ответа
на учебное или экзаменационное задание.
Издательской группы
'Гранд-Фаир'
можно приобрести ------ в московских магазинах: -------
1. Московский Дом Книги
Адрес: г.
Москва, ул. Новый Арбат, д. 8 (м. Арбатская). Справочные тел.: 290-35-80,
290-45-07
2. Торговый Дом 'Москва'
Адрес: г.
Москва, ул. Тверская, д. 8 (м. Тверская). Справочный тел.: 229-64-83
3. 'Библио-Глобус'
Адрес: г.
Москва, ул. Мясницкая, д. 6 (м. Лубянка). Справочный тел.: 928-35-67
4. 'Молодая гвардия'
Адрес: г. Москва,
ул. Б. Полянка, д. 28 (м. Полянка).
Справочный тел.: 238-50-01
5. 'Белые облака'
Адрес: г.
Москва, ул. Покровка, д. 4. Справочный
тел.: 921-61-25
6. Дом Педагогической Книги
Адрес: г.
Москва, ул. Б. Дмитровка, д. 7/5. Справочные тел.: 229-50-04, 229-93-42
7. Дом Книги в Медведково
Адрес: г.
Москва, Заревый пр-д, д. 12 (м. Медведково). Справочный тел.: 478-48-97
8. Дом Медицинской Книги
Адрес: г.
Москва, Комсомольский просп., д. 25 (м. Фрунзенская). Справочные тел.: 245-39-33,
245-39-27, 248-28-46
9. Дом Технической Книги
Адрес: г.
Москва, Ленинский просп.,
д. 40. Справочные тел.: 137-68-88, 137-60-19 10.
'Путь к себе'
Адрес: г.
Москва, Ленинградский просп., д. 10а (м. Белорусская). Справочный тел.: 257-39-87
Издательская группа 'ГРАНД-ФАИР'
приглашает к сотрудничеству авторов и книготорговые
организации
Телефон / факс: (095)721 - 38
- 56
(многоканальный)
Почтовый адрес:
109428, Москва, ул. Зарайская, д. 47, корп. 2
e-mail:
office@grand-fair.ru
Интернет: http://www.grand-fair.ru
Серия 'Наука & Жизнь'
Бартон Гуттман, Энтони Гриффитс, Дэвид Сузуки, Тара
Куллис
ГЕНЕТИКА
Дизайн обложки
Е. Ярошенко
ЛР 065864 от 30
апреля 1998 г.
Подписано в
печать 20.09.2004.
Формат 84 х 108 1/32.
Бумага офсетная.
Гарнитура
'Таймс'. Печать офсетная.
Усл. печ. л.
23,52. Тираж 3000 экз.
Заказ 4707.
Издательство
'ФАИР-ПРЕСС' 109428, Москва, ул. Зарайская, д. 47, корп. 2
Отпечатано в
полном соответствии
с качеством предоставленных
диапозитивов
в ОАО 'Можайский
полиграфический комбинат'
143200, г. Можайск, ул. Мира, 93
Сканирование
и форматирование: Янко Слава (Библиотека
Fort/Da) || slavaaa@yandex.ru || yanko_slava@yahoo.com || http://yanko.lib.ru || Icq# 75088656 || Библиотека: http://yanko.lib.ru/gum.html
|| Номера страниц - внизу. АНОНС КНИГИ