


50-51
Биомолекулы. Углеводы
Гликозаминогликаны и гликопротеины
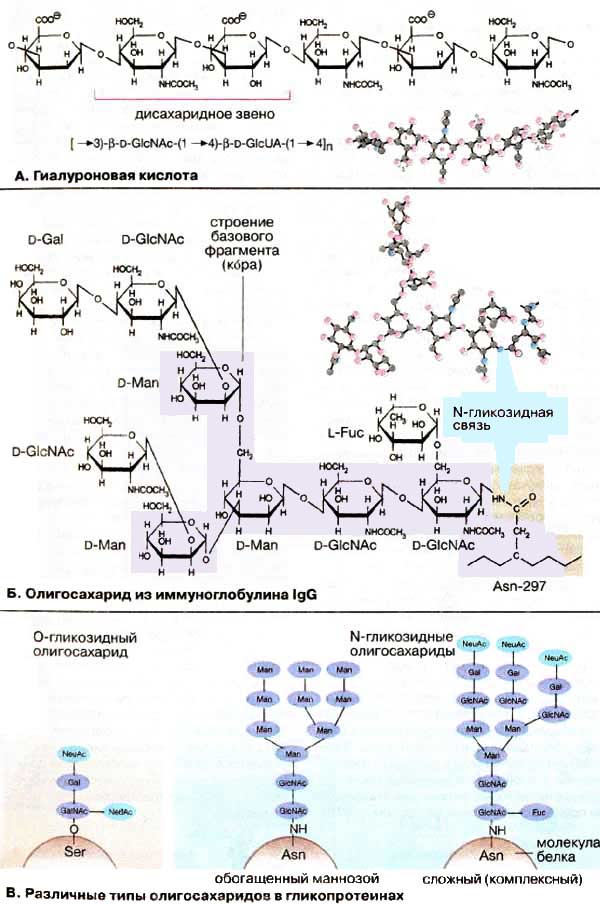 А.Гиалуроновая кислота
А.Гиалуроновая кислота
Гликозаминогликаны, группа кислых
гетерополисахаридов, в качестве структурных элементов протеогликанов
являются важным компонентом межклеточного матрикса (см. с. 336).
В качестве типовых структурных блоков
гликозаминогликаны содержат аминосахара, такие, как
глюкуроновая или идуроновая кислоты. Большинство полисахаридов этой группы в
различной степени этерифицировано остатками серной кислоты, которые усиливают их
кислотные свойства. Гликозаминогликаны присутствуют в организме позвоночных как
в свободном виде, так и в составе протеогликанов.
Гиалуроновая кислота, относительно
простой неэтерифицированный гликозаминогликан, построена из дисахаридных
звеньев, состоящих из N-ацетилглюкозамина и глюкуроновой кислоты,
соединенных в положении β(1→3). Повторяющиеся звенья связаны в
положении β(1→4). Благодаря присутствию β(1→3)-cвязeй молекула гиалуроновой
кислоты, насчитывающая несколько тысяч моносахаридных остатков, принимает
конформацию спирали. На один виток спирали приходится три дисахаридных блока.
Локализованные на внешней стороне спирали гидрофильные карбоксильные группы
остатков глюкуроновой кислоты могут связывать ионы Ca2+. За счет
сильной гидратация этих групп гиалуроновая кислота и другие
Гликозаминогликаны при образовании гелей связывают 10 000-кратный объем воды.
Гиалуроновая кислота выполняет функцию стабилизатора геля в стекловидном теле
глаза, которое содержит всего 1% гиалуроновой кислоты и на 98% состоит из
воды.
Б. Олигосахарид из иммуноглобулина IgG
Многие белки внешней стороны плазматических
мембран и большинство секретируемых белков содержат олигосахаридные цепи,
которые синтезируются в процессе посттрансляционной модификации в
эндоплазматическом ретикулуме и в аппарате Гольджи (см. с. 226).
Цитоплазматические белки, напротив, редко бывают
гликозилированы. Гликопротеины могут содержать до 50% углеводов, но, как
правило, в молекуле преобладает белковая часть.
В качестве примера на схеме представлена
структура олигосахарида имммуноглобулина IgG (см. с. 256). Олисахарид связан
Ν-гликозидной связью с амидной группой остатка аспарагина в
Fс-области тяжелой цепи белка. Функция олигосахарида не
установлена.
В молекуле олигасахарида имеется Т-образный
базовый фрагмент (кор) из двух остатков Ν-ацетилглюкозамина и трех
остатков маннозы (на схеме фиолетового цвета). Наличие такого фрагмента
характерно для всех Ν-гликозидных олигосахаридов. Кроме того, в молекуле
содержится еще два остатка Ν-ацетиглюкозамина, по одному остатку фукозы и
галактозы. В гликопротеинах встречаются самые разные типы ветвлений. В
приведенной структуре наряду со связью β(1→4) имеется связь β(1→2), а также
мостики в положении α(1→3) и α(1→6).
В. Различные типы олигосахаридов в
гликопротеинах
В некоторых гликопротеинах наряду с
N-гликозидными олигосахаридами (по остатку аспарагина) встречаются, хотя и не
столь часто, олигосахариды, связанные О-гликозидной связью (с
гидроксильной группой остатков серина и треонина).
Различают два типа N - гликозидсвязанных
олигосахаридов, которые синтезируются по различным механизмам. При
гликозилировании в эндоплазматическом ретикулуме к апобелку вначале
присоединяется олигосахарид, включающий приведенный на схеме кор с шестью
дополнительными остатками маннозы и тремя концевыми остатками глюкозы (см. с.
226). При отщеплении от первичного олигосахарида остатков глюкозы образуется
простая форма олигосахарида (обогащенного остатками маннозы). При этом к
олигосахариду не присоединяются другие типы моносахаридов. Отщепление остатков
маннозы с заменой на другие моносахариды приводит к образованию сложного
(комплексного) олигосахарида, который представлен на схеме справа. В
комплексных олигосахаридах часто содержатся концевые остатки
N-ацетилнейраминовой кислоты, которые придают им отрицательный
заряд.