


186-187
Метаболизм белков
Биосинтез аминокислот
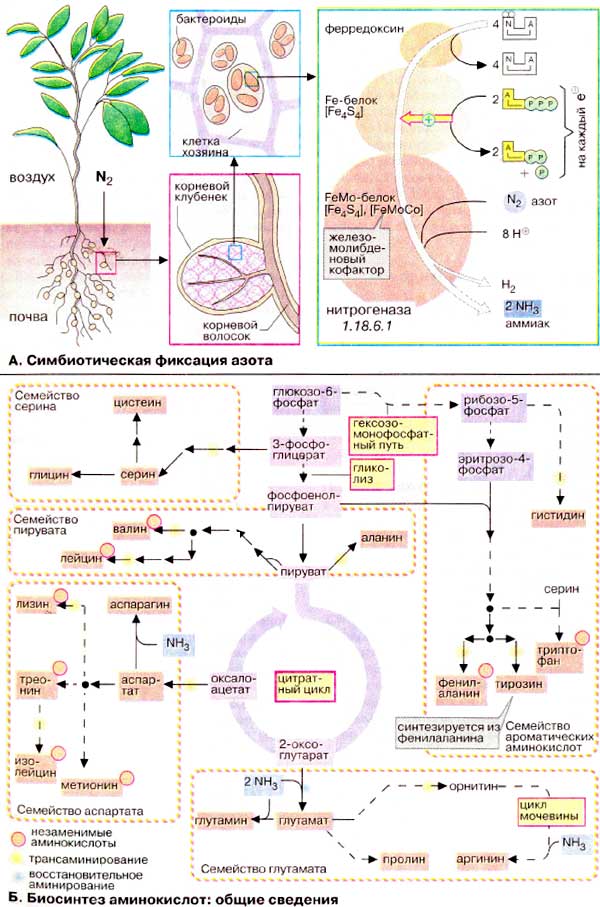 В атмосфере элементарный азот
(Ν2) присутствует практически в неограниченном количестве. Прежде чем
поступить в круговорот азота, он должен быть восстановлен до NH3 и
включен ('фиксирован') в аминокислоты.
В атмосфере элементарный азот
(Ν2) присутствует практически в неограниченном количестве. Прежде чем
поступить в круговорот азота, он должен быть восстановлен до NH3 и
включен ('фиксирован') в аминокислоты.
А. Симбиотическая фиксация
азота
Фиксировать атмосферный азот способны
лишь немногие виды бактерий и синезеленых водорослей. Они находятся в почве
свободно или живут в симбиозе с растениями. Особо важное хозяйственное значение
имеет симбиоз между бактериями рода Rhizobium и бобовыми
растениями (Fabales), такими, как клевер, бобы или горох. Эти растения
очень питательны благодаря высокому содержанию белка.
В симбиозе с бобовыми бактерии живут в
корневых клубочках внутри растительных клеток, так называемые
бактероидах. С одной стороны, растение снабжает бактериоды питательными
веществами, а с другой, извлекает пользу от фиксированного азота, который
поставляет симбионт. Фиксирующим N2 ферментом бактерий является
нитрогеназа. Она состоит из двух компонентов: Fe-белка и FeMo-белка.
Fe-белок, содержащий [FeS4]-центр (см. с. 144), служит
окислительно-восстановительной системой, которая принимает электроны от
ферредоксина и передает их во второй компонент, FeMo-белок. Этот
молибденсодержащий белок переносит электроны на N2 и таким образом
через различные промежуточные стадии продуцирует NH3. Часть
восстановительных эквивалентов переносится в побочной реакции на H+.
Поэтому наряду с NH3 всегда образуется водород.
Б. Биосинтез аминокислот: общие
сведения
По особенностям биосинтеза протеиногенные аминокислоты (см.
с. 66) подразделяются на пять семейств. Члены каждого семейства
имеют общих предшественников, которые образуются в цитратном цикле или при катаболизме
углеводов. Пути биосинтеза здесь приведены схематически, более подробно они
рассматриваются на сс. 400 и 401.
В то время как растения и микроорганизмы
могут вполне синтезировать все аминокислоты, млекопитающие в ходе эволюции
утратили способность к синтезу примерно половины из 20 протеиногенных
аминокислот. Поэтому незаменимые аминокислоты должны поступать с пищей.
Так, организм высших организмов не способен синтезировать ароматические
аминокислоты de novo (тирозин не является незаменимой аминокислотой только
потому, что может образоваться из фенилаланина). К незаменимым аминокислотам
принадлежат аминокислоты с разветвленной боковой цепью: валин и
изолейцин, а также лейцин, треонин, метионин и лизин. Гистидин и аргинин
являются незаменимыми для крыс, но касается ли это также человека - спорно.
Наличие незаменимых аминокислот в рационе питания, по-видимому, существенно по
крайней мере во время роста организма. Питательная ценность белков (см. с. 348)
решающим образом зависит от содержания незаменимых аминокислот. Растительные
белки зачастую бедны лизином или метионином. В то же время животных белки
содержат все аминокислоты в сбалансированных соотношениях.
Заменимые аминокислоты (аланин,
аспарагиновая и глутаминовая кислоты и их амиды, аспарагин и глутамин)
образуются в результате трансаминирования из промежуточных метаболитов -
2-кетокислот. Пролин синтезируется в достаточных количествах из глутамата, а
представители серинового семейства (серин, глицин и цистеин) сами являются
естественными метаболитами организма животных.