


302-303
Ткани и органы. Печень
Метаболизм углеводов
 Глюкоза, наряду с жирными кислотами и
кетоновыми тепами, является важнейшим источников энергии. Уровень глюкозы
в крови поддерживается постоянным 4-6 мМ (0,8-1,0 г/л) благодаря тонкой
регуляции процессов ее поступления и потребления. Глюкоза поступает из кишечника
(за счет переваривания пищи), печени и почек. При этом печень выполняет функцию
'глюкостата': в фазе резорбции глюкоза поступает в печень из крови и
накапливается в виде гликогена. При дефиците глюкозы (фаза пострезорбции,
голодание) печень, напротив, поставляет глюкозу, которая образуется за счет
процессов гликогенолиза и глюконеогенеза (см. с. 300).
Глюкоза, наряду с жирными кислотами и
кетоновыми тепами, является важнейшим источников энергии. Уровень глюкозы
в крови поддерживается постоянным 4-6 мМ (0,8-1,0 г/л) благодаря тонкой
регуляции процессов ее поступления и потребления. Глюкоза поступает из кишечника
(за счет переваривания пищи), печени и почек. При этом печень выполняет функцию
'глюкостата': в фазе резорбции глюкоза поступает в печень из крови и
накапливается в виде гликогена. При дефиците глюкозы (фаза пострезорбции,
голодание) печень, напротив, поставляет глюкозу, которая образуется за счет
процессов гликогенолиза и глюконеогенеза (см. с. 300).
Печень обладает свойством синтезировать
глюкозу из других сахаров, например фруктозы и галактозы, или из
других продуктов промежуточного метаболизма. Превращение лактата в глюкозу в
цикле Кори (см. с. 330) и аланина в глюкозу в цикле аланина (см.
с. 330) играет особую роль в обеспечении эритроцитов и мышечных
клеток.
Необходимыми условиями активного
углеводного обмена в печени является обратимый транспорт сахаров через
плазматическую мембрану гепатоцитов (при отсутствии контроля инсулином) и
наличие фермента глюкозо-6-фосфатазы, высвобождающего глюкозу из
глюкозо-6-фосфата.
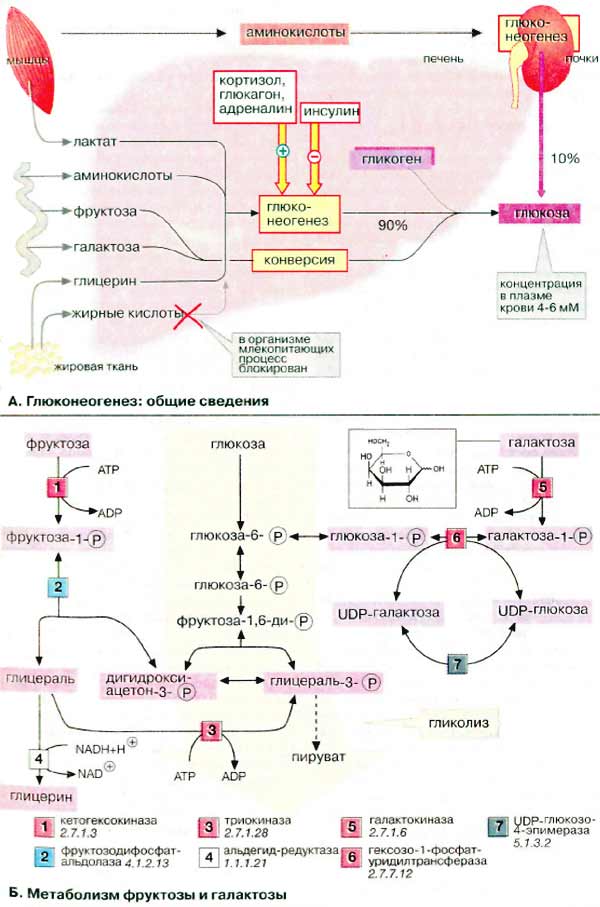
А. Глюконеогенез: общие
сведения
Синтез глюкозы de novo (до 250 г
в сутки) происходит в основном в печени. Процесс глюконеогенеза может идти и в
почках, однако из-за небольших размеров почек их вклад в синтез глюкозы
составляет всего 10%.
Глюконеогенез контролируется гормонами.
Кортизол, глюкагон и адреналин стимулируют этот процесс, а
инсулин, напротив, подавляет.
При глюконеогенезе в печени наиболее
важными субстратами являются лактат, поступающий из мышечной ткани и
эритроцитов, аминокислоты из желудочно-кишечного тракта (глюкогенные
аминокислоты) и мышц (аланин), а также глицерин из жировых тканей. В
почках в качестве субстрата служат главным образом аминокислоты (см. с.
320).
Жирные кислоты и другие источники
ацетил-КоА не могут использоваться в организме млекопитающих для биосинтеза
глюкозы, поскольку ацетил-КоА, образующийся при β-окислении в цитратном цикле
(см. с. 140), полностью окисляется до СО2, в то время как в
глюконеогенезе исходным продуктом является оксалоцетат.
Б. Метаболизм фруктозы и
галактозы
Метаболизм фруктозы осуществляется
превращением ее в глюкозу (на схеме слева). Вначале фруктоза фосфорилируется при
участии фермента кетогексокиназы (фруктокиназы) [1] с образованием
фруктозо-1-фосфата, который далее расщепляется альдолазой до глицеральдегида
(глицераля) и дигидроксиацетон-3-фосфата [2]. Последний уже является
промежуточным продуктом гликолиза (в центре схемы), а глицераль фосфорилируется
в присутствии триокиназы, образуя глицераль-3-фосфат [3].
Затем глицеральдегид частично восстанавливается до глицерина
[4] или окисляется до глицерата. После фосфорилирования
оба соединения вновь включаются в гликолиз (на схеме не приведено). При восстановлении
глицеральдегида [4]расходуется НАДН (NADH). Поскольку
при конверсии этанола лимитирующим фактором является низкое соотношение концентраций
НАД+/НАДН (NAD+/NADH). Этот процесс ускоряется в присутствии
фруктозы (см. с. 312).
Кроме того, в печени реализуется
полиольный путь трансформации фруктозы в глюкозу (на схеме не приведен):
фруктоза за счет восстановления С-2 превращается в сорбит, а при последующем
дегидрировании С-1 - в глюкозу.
Метаболизм галактозы также начинается с фосфорилирования
с образованием галактозо-1-фосфата [5] (на схеме
справа). Далее следует эпимеризация С-4 с образованием производного глюкозы.
Биосинтез УДФ-глюкозы (UDP-глюкозы), промежуточного продукта обмена глюкозы,
осуществляется обходным путем - через УДФ-галактозу (UDP-галактозу) и последующую
эпимеризацию [6, 7]. По такому же пути идет биосинтез
самой галактозы, поскольку все реакции за исключением [5]
обратимы.