


330-331
Ткани и органы. Мышцы
Метаболическая регуляция мышечного сокращения
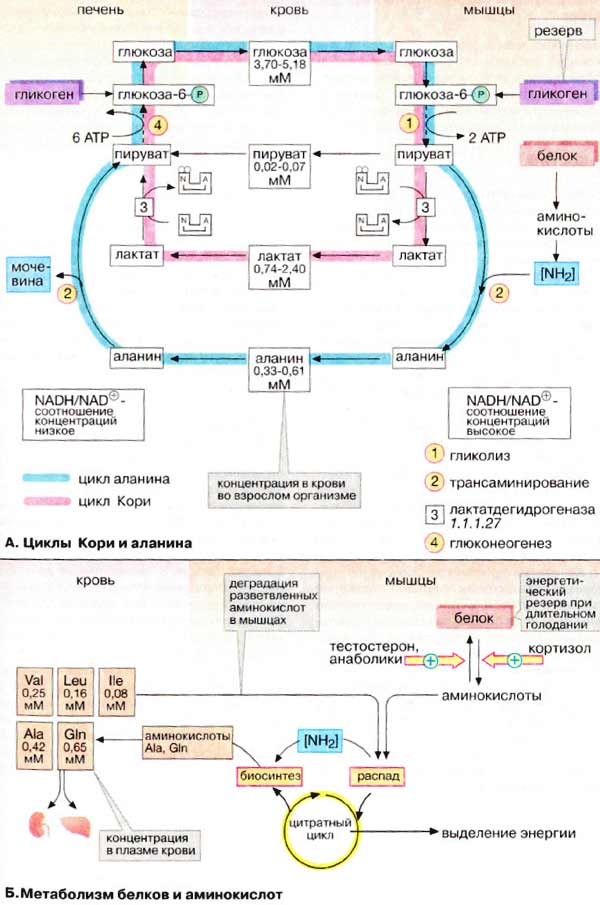 А. Циклы Кори и
аланина
А. Циклы Кори и
аланина
В клетках, не содержащих митохондрий
(например, в эритроцитах), или в тканях при недостаточном снабжении кислородом
(например, в активно работающих мышцах) АТФ (АТР) синтезируется за счет процесса
превращений глюкозы в лактат, т. е. за счет процесса брожения (= анаэробного
гликолиза, см. рис. 153). Лактат переносится кровью в печень, где в
процессе глюконеогенеза с затратой АТФ (см. рис. 157) вновь конвертируется
в глюкозу (цикл Кори).
При интенсивной работе мышцы максимально
активируется гликолиз. Продукт гликолиза, пировиноградная кислота (пируват)
накапливается в цитоплазме и недостаточно быстро поступает в митохондрии, если
они из-за недостатка кислорода не готовы к окислению пирувата. В анаэробных
условиях пируват в реакции, катализируемой лактатдегидрогеназой (см. рис.
103) (заключительный этап гликолиза) восстанавливается до лактата.
Одновременно
НАДН (NADH), кофермент лактатдегидрогеназы, окисляется до НАД+
(NAD+), который вновь используется на окислительном этапе
гликолитического пути. Этой реакции способствует относительно высокое отношение
НАДН/НАД+ в мышечной ткани. Лактат диффундирует в кровь и поступает в
печень, где конвертируется в глюкозу. Таким образом, образование лактата
временно заменяет аэробный метаболизм глюкозы и частично переносит этот процесс
из мышц в печень.
Обратная связь, подобная циклу Кори,
существует в цикле аланина, в котором также участвует пируват.
Цикл аланина берет начало с протеолиза белков. Образующиеся аминокислоты
в результате трансаминирования в присутствии ферментов превращаются в
α-кетокислоты (на схеме не приведено, см. рис. 181), которые в основном включаются
в цикл трикарбоновых кислот (цитратный цикл) (см. рис. 183).Одновременно в
реакции, катализируемой аланинтрансаминазой, аминогруппы из разных аминокислот
переносятся на имеющийся субстрат, пируват. Образующийся аланин поступает
в кровь и переносится в печень. Таким образом, цикл аланина служит каналом
передачи азота и предшественников глюкозы в печень, которая является местом
синтеза конечных продуктов азотистого обмена, например мочевины (см. рис.
185).
Следует напомнить, что при анаэробном
гликолизе образуются кислоты, которые, не принимая участие в последующем обмене,
существуют в форме анионов. Поэтому при интенсивном анаэробном гликолизе рН
мышечной клетки может понизиться настолько, что сокращение станет невозможным.
Обычно этого не происходит благодаря быстрому выходу кислых метаболитов (лактата
и пирувата) в кровь, которая также может оказаться закисленной
(метаболический ацидоз).
Б. Метаболизм белков и
аминокислот
Скелетные мышцы активно участвуют в
метаболизме аминокислот. Это наиболее важный участок деградации разветвленных
аминокислот (Val и Ile, см. с. 402). Ряд других аминокислот также
деградируются преимущественно в мышцах. Одновременно идет ресинтез и
высвобождение в кровь аланина и глутамина. Эти аминокислоты служат
переносчиками азота, образующегося при расщеплении белков, в печень (цикл
аланина) и почки (см. рис. 319).
При голодании мышечные белки
служат энергетическим резервом организма. Они гидролизуются до аминокислот,
которые поступают в печень. Здесь углеродный скелет аминокислот конвертируется в
промежуточные продукты цитратного цикла, в том числе в ацетоацетил-КоА и
ацетил-КоА (см. рис. 183). Эти амфиболические соединения окисляются в цикле
трикарбоновых кислот или включаются в процесс
глюконеогенеза.
Синтез и расщепление мышечных белков
контролируются гормонами. Тестостерон и синтетические анаболики
стимулируют биосинтез белка; напротив, кортизол подавляет образование
мышечных белков.
Белки актин и миозин содержат остатки
гистидина, метилированного на стадии посттрансляционной модификации. При
расщеплении этих белков образуется 3-метилгистидин, который дальше не
разрушается. Количество метилгистидина в моче служит мерой деградации мышечных
белков.