


326-327
Ткани и органы. Мышцы
Регуляция сокращения мышечных волокон
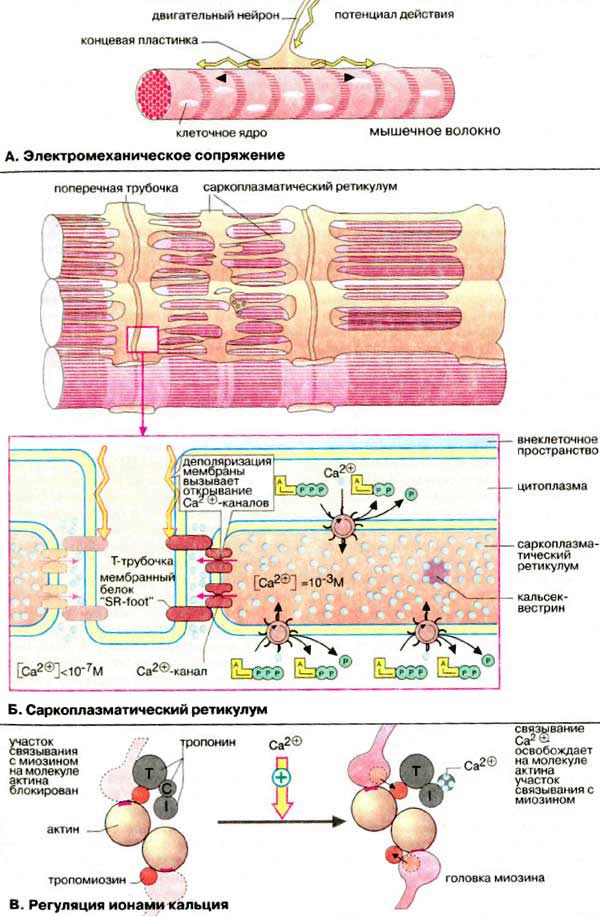 А. Электромеханическое
сопряжение
А. Электромеханическое
сопряжение
Сокращением мышечного волокна управляют двигательные
нейроны, которые выделяют нейромедиатор ацетилхолин в нервно-мышечные
соединении (синапсы) (см. рис. 345). Ацетилхолин диффундирует
через синаптическую щель и взаимодействуют с ацетилхолиновыми (холинэргическими)
рецепторами плазматической мембраны мышечных клеток. Это вызывает открывание
трансмембранных ионных каналов и деполяризацию клеточной мембраны (образование
потенциала действия). Потенциал действия быстро распространяется по всем
направлениям от нервно-мышечного соединения (см. рис. 341,
343), возбуждая все мышечные клетки. В течение нескольких
миллисекунд реализуется рассмотренный выше цикл сокращения мышечного волокна.
Б. Саркоплазматический
ретикулум
Саркоплазматический ретикулум [СР (SR)]
-разветвленная подобная эндоплазматическому ретикулуму органелла, окружающая
индивидуальные миофибриллы подобно сетке (в верхней части схемы в качестве
примера приведен СР клетки сердечной мышцы). В покоящихся клетках концентрация
Са2+ очень низка (менее 10-5 М). Однако в
саркоплазматическом ретикулуме уровень ионов Са2+ существенно
выше (около 10-3 М). Высокая концентрация Са2+ в СР
поддерживается Са2+-АТФ-азами. Кроме того, в СР имеется
специальный белок кальсеквестрин (55 кДа), который благодаря высокому
содержанию кислых аминокислот способен прочно связывать ионы
Са2+.
Переносу потенциала действия на СР
индивидуальной миофибриллы способствуют поперечные трубочки Т-системы,
представляющие трубчатые впячивания клеточной мембраны и находящиеся в тесном
контакте с индивидуальными миофибриллами. Деполяризация плазматической мембраны
передается через Т-трубочки на потенциал-управляемый мембранный
белок (так называемый "SR-foot") прилегающей мембраны СР, который открывает
Са2+-каналы. Результатом является выброс ионов Са2+
из СР в пространство между филаментами актина и миозина до уровня
≥10-5 M. В конечном итоге выброс ионов Са2+ является
пусковым механизмом процесса сокращения миофибрилл.
В. Регуляция ионами
кальция
В расслабленной скелетной мышце комплекс
тропонина (субъединицы = Т, С, I) с тропомиозином
препятствует взаимодействию миозиновых головок с
актином.
Быстрое увеличение в цитоплазме
концентрации ионов кальция в результате открывания каналов СР приводит к
связыванию Са2+ с С-субъединицей тропонина. Последняя по свойствам
близка кальмодулину (см. рис. 375). Связывание ионов Са2+ вызывает
конформационную перестройку в тропонине, тропонинтропомиозиновый комплекс
разрушается и освобождает на молекуле актина участок связывания с миозином (на
схеме выделен красным цветом). Это инициирует цикл мышечного сокращения (см. с.
324)
В отсутствие последующего стимулирования
АТФ-зависимые кальциевые насосы мембраны СР быстро снижают концентрацию ионов
Ca2+ до исходного уровня. Как следствие, комплекс Са2+ с
тропонином С диссоциирует, тропонин восстанавливает исходную конформацию, место
связывания миозина на актине блокируется и мышца расслабляется.
Таким образом, при сокращении
мышечного волокна скелетных мышц позвоночных происходит следующая
последовательность событий. При поступлении сигнала от двигательного нейрона
мембрана мышечной клетки деполяризуется, сигнал передается на
Сa2+-каналы СР. Са2+-каналы открываются, внутриклеточный
уровень ионов Са2+ возрастает. Ионы Сa2+ связывается с
тропонином С, вызывая конформационную перестройку в тропонине, что влечет за
собой разрушение комплекса тропонин-тропомиозин и дает возможность головкам
миозина связываться с актином. Происходит инициация актин-миозинового
цикла.
По завершении сокращения уровень
ионов Са2+ снижается за счет активного обратного транспорта
Са2+ в СР, тропонин С отдает Са2+, комплекс
тропонин-тропомиозин занимает исходное положение на молекуле актина, блокируя
актин-миозиновый цикл. Результатом является расслабление
мышцы.