


328-329
Ткани и органы. Мышцы
Источники энергии
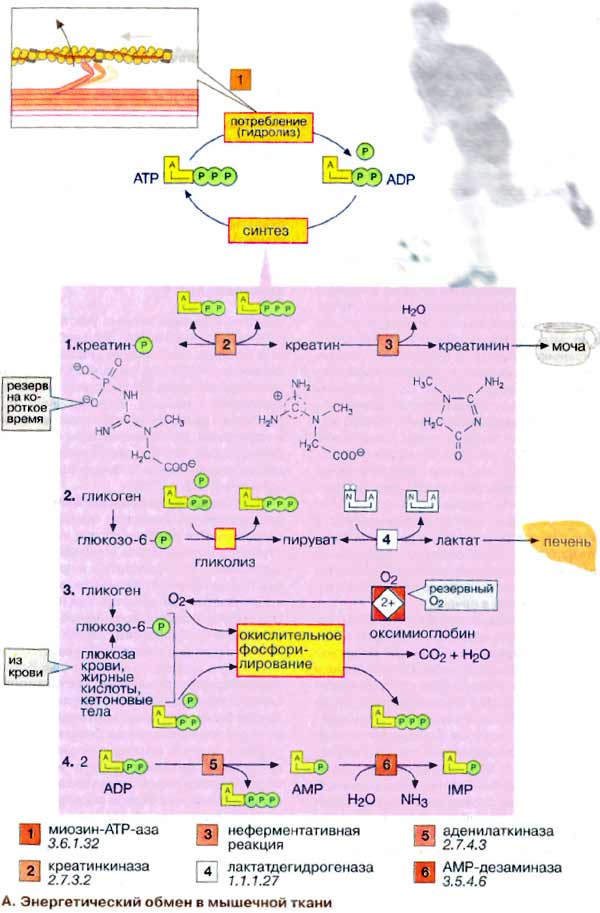 А. Энергетический обмен в мышечной
ткани
А. Энергетический обмен в мышечной
ткани
Важнейшей функцией мышечного волокна
является сократительная. Процесс сокращения и расслабления связан с
потреблением АТФ (АТР), гидролиз которого катализирует миозин-АТФ-аза
[1] (см. рис. 325). Однако небольшой запас АТФ, имеющийся в мышцах,
расходуется менее чем за 1 с после стимуляции.
Потребности работающей мышцы в
АТФ удовлетворяются за счет следующих ферментативных реакций:
1. Резерв в виде креатинфосфата.
Быстрая регенерация АТФ может быть достигнута за счет переноса фосфатной группы
креатинфосфата на АДФ (ADP) в реакции, катализируемой креатинкиназой [2].
Однако и этот мышечный резерв 'высокоэргического фосфата' расходуется в
течение нескольких секунд. В спокойном состоянии креатинфосфат вновь
синтезируется из креатина. При этом фосфатная группа присоединяется по
гуанидиновой группе креатина (N-гуанидино-N-метилглицина). Креатин,
который синтезируется в печени, поджелудочной железе и почках, в основном
накапливается в мышцах. Здесь креатин медленно циклизуется за счет
неферментативной реакции [3] с образованием креатинина, который поступает
в почки и удаляется из организма (см. рис. 317).
2 Анаэробный гликолиз. В мышечной
ткани наиболее важным долгосрочным энергетическим резервом является
гликоген (см. рис. 159). В покоящейся ткани содержание гликогена составляет
до 2% от мышечной массы. При деградации под действием фосфорилазы
гликоген легко расщепляется с образованием глюкозо-6-фосфата, который при
последующем гликолизе превращается в пируват. При большой потребности в АТФ и
недостаточном поступлении кислорода пируват за счет анаэробного гликолиза
восстанавливается в молочную кислоту (лактат), которая диффундирует в
кровь (цикл Кори, см. рис. 331).
3. Окислительное
фосфорилирование. В аэробных условиях образующийся пируват поступает в
митохондрии, где подвергается окислению. Окислительное фосфорилирование (см. с.
143) - наиболее эффективный и постоянно действующий путь синтеза АТФ. Однако
этот путь реализуется при условии хорошего снабжения мышц кислородом. Наряду с
глюкозой, образующейся при расщеплении мышечного гликогена, для синтеза АТФ
используются и другие "энергоносители", присутствующие в крови: глюкоза
крови, жирные кислоты и кетоновые тела.
4. Образование инозинмонофосфата
[ИМФ (IMP)]. Другим источником быстрого восстановления уровня АТФ является
конверсия АДФ в АТФ и АМФ (AMP), катализируемая аденилаткиназой (миокиназой)
[5]. Образовавшийся АМФ за счет дезаминирования частично превращается
в ИМФ (инозинмонофосфат) (см. рис. 191), что сдвигает реакцию в нужном
направлении.
Из всех способов синтеза АТФ наиболее
продуктивным является окислительное фосфорилирование. За счет этого
процесса обеспечиваются потребности в АТФ постоянно работающей сердечной мышцы
(миокарда). Вот почему для успешной работы сердечной мышцы обязательным условием
является достаточное снабжение кислородом (инфаркт миокарда - это
следствие перебоев в поступлении кислорода).
В высокоактивных (красных) скелетных
мышцах источником энергии для рефосфорилирования АДФ служит окислительное
фосфорилирование в митохондриях. В обеспечении этих мышц кислородом принимает
участие миоглобин (Mb) - близкий гемоглобину белок, обладающий свойством
запасать кислород. В малоактивных скелетных мышцах, лишенных красного миоглобина
и поэтому белых, главным источником энергии для восстановления уровня АТФ
является анаэробный гликолиз. Такие мышцы сохраняют способность к быстрым
сокращениям, однако они могут работать лишь короткое время, поскольку при
гликолизе образование АТФ идет с низким выходом. Спустя некоторое время мышцы
истощаются в результате изменения рН в мышечных клетках.
Расщепление гликогена контролируется
гормонами (см. рис. 123). Процесс гликогенолиза стимулируется
адреналином (через b-рецепторы) за счет образования цАМФ и активации
киназы фосфорилазы. Активация фосфорилазы наступает также при увеличении
концентрации ионов Са2+ во время мышечного
сокращения.