


380-381
Рост и развитие. Деление клеток
Клеточный цикл
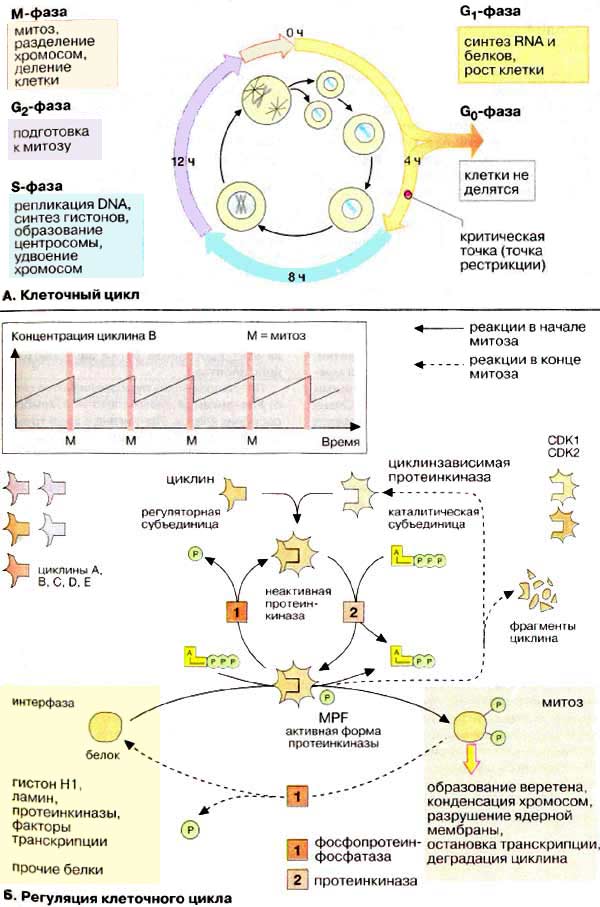 А. Клеточный цикл
А. Клеточный цикл
Характерным свойством пролиферирующих
клеток является их способность к делению.
У животных клеток интервал между митозами (клеточный цикл,
точнее митотический цикл) составляет примерно 10-24 ч (в примере, приведенном
на схеме, 24 ч). За это время клетка проходит четыре фазы жизненного цикла:
G1-фазу начального роста, S-фазу удвоения молекул ДНК
(репликации, см. рис. 239), G2-фазу
роста и М-фазу клеточного деления. Наиболее детально изучена фаза клеточного
деления, митоз (М-фаза). В G1-фазе, продолжительность которой может
сильно варьировать, происходит синтез мРНК, белков и других компонентов клетки.
У некоторых клеток в жизненном цикле может отсутствовать G1-фаза.
Клетки, которые прошли дифференцировку и больше не делятся, постоянно находятся
в фазе покоя G0 . При стимуляции митогенами (например, ростовыми
факторами, онкогенными вирусами) покоящиеся клетки могут вернуться в состояние,
свойственное фазе G1. ЕСЛИ такие клетки пройдут критическую точку,
они вступают в S-фазу. G2-фаза является конечным этапом подготовки
клетки к делению.
В совокупности фазы G1,
G0, S и G2 носят название интерфазы. В клеточном
цикле интерфаза сменяется существенно более короткой фазой митоза
(М).
Б. Регуляция клеточного
цикла
Регуляция клеточного цикла
осуществляется посредством обратимого фосфорилирования/дефосфорилирования
регуляторных белков (см. рис. 117). Ключевым белком, регулирующим вступление
клетки в митоз (G2/M-переход), является специфическая
серин/треонин-протеинкиназа, которая носит название фактор созревания [ФС
(MPF, от англ. maturation promoting factor)]. В активной
форме фермент катализирует фосфорилирование многих белков, принимающих участие в
митозе, таких, например, как входящий в состав хроматина гистон H1 (см.
с. 236), ламин (компонент цитоскелета, обнаруженный в ядерной мембране),
факторы транскрипции, белки митотического веретена и ряд ферментов.
Фосфорилирование этих белков запускает процесс митоза. После завершения
митоза регуляторная субъединица ФС, циклин, маркируется убиквитином и
подвергается протеолизу (см. с. 178). Теперь наступает очередь протеинфосфатаз,
которые дефосфори пируют белки, принимавшие участие в митозе, после чего клетка
возвращается в состояние интерфазы.
ФС - гетеродимерный фермент,
включающий регуляторную субъединицу, циклин, и каталитическую
субъединицу, циклинзависимую киназу [ЦЗК (CDK от англ. cyclin
dependent kinase) или p34cdc2; 34 кДа]. Активной формой
фермента является лишь димер ЦЗК+циклин. Кроме того, активность протеинкиназы
регулируется путем обратимого фосфорилирования самого фермента (на схеме
представлен предельно простой вариант этого процесса).
В клетках позвоночных присутствует ряд
различных циклинов и циклинзависимых киназ. Разнообразные
сочетания двух субъединиц фермента регулируют запуск митоза, начало процесса
транскрипции в G1-фазе, переход критической точки после завершения
транскрипции, начало процесса репликации ДНК в S-периоде интерфазы (стартовый
переход) и другие ключевые переходы клеточного цикла (на схеме не
приведены).
В ооцитах лягушки вступление в митоз
(G2/M-переход) регулируется путем изменения концентрации циклина.
Циклин непрерывно синтезируется в интерфазе до достижения максимальной
концентрации в фазе М, когда запускается весь каскад фосфорилирования белков,
катализируемый ФС. К окончанию митоза циклин быстро разрушается протеиназами,
также активируемыми ФС. В других клеточных системах активность ФС регулируется
за счет различной степени фосфорилирования самого
фермента.