


126-127
Метаболизм. Энергетика
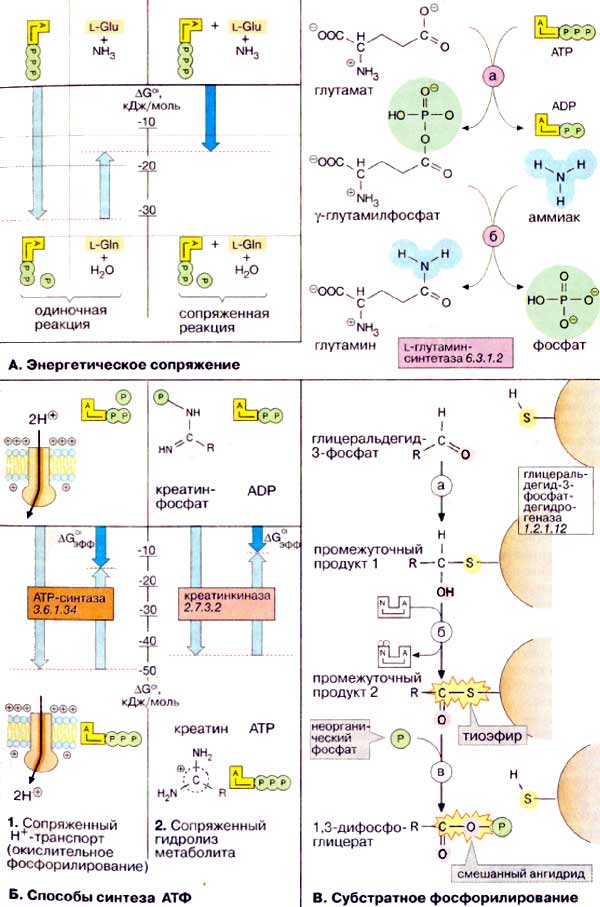
Энергетическое сопряжение
 В клетке химическая энергия запасается в
виде так называемых 'высокоэнергетических' метаболитов. Наиболее важным таким
метаболитом, макроэргом (см. с. 22), обеспечивающим энергией большое
число энергозависимых реакций, является АТФ.
В клетке химическая энергия запасается в
виде так называемых 'высокоэнергетических' метаболитов. Наиболее важным таким
метаболитом, макроэргом (см. с. 22), обеспечивающим энергией большое
число энергозависимых реакций, является АТФ.
А. Энергетическое сопряжение
В качестве меры потенциала переноса фосфатных групп у высокоэнергетических
соединений произвольно выбрано изменение свободной энергии гидролиза ΔGo'
(см. сс. 24, 124). Это, однако, не
означает, что АТФ (АТР) в энергетически сопряженных реакциях будет действительно
гидролизоваться. Гидролиз АТФ без сопряжения с эндергоническим процессом приводит
лишь к выделению тепла. Сопряжение двух реакций возможно при наличии общего
промежуточного продукта. Поясним это положение на примере реакции с участием
глутаминсинтетазы. Сначала концевая фосфатная группа переносится с АТФ
на глутамат с образованием высокоэнергетического смешанного ангидрида
(а). На втором этапе (б)
фосфатная группа промежуточного продукта вытесняется NH3 с образованием
глутамина и свободного фосфата. Баланс и величина ΔGo' суммарной
реакции соответствуют сумме балансов и значений свободных энергий отдельных
реакций.
Б. Способы синтеза АТФ
Так как синтез АТФ является
высоко эндоэргической реакцией, он должен сопрягаться с другим высоко
экзоэргическим процессом. В ходе эволюции сформировались два важных способа
синтеза АТФ, которые реализуются по всех клетках. Наиболее эффективный способ синтеза АТФ
использует энергию градиента электрохимического потенциала (см. с. 128)
для образования АТФ из АДФ (ADP) и неорганического фосфата. Энергия для создания
такого градиента возникает в результате окислительно-восстановительного
процесса. Этот механизм называют окислительным фосфорилированием.
Транспортирующая H+ АТФ-синтаза
(см. с. 144) использует для синтеза АТФ энергию градиента потенциала. У
эукариот окислительное фосфорилирование происходит только в присутствии
кислорода (т. е. в аэробных условиях).
Второй, эволюционно более ранний способ синтеза АТФ осуществляется в анаэробных условиях. Он основан на переносе фосфатных остатков на АДФ через метаболит с высоким потенциалом переноса фосфатных групп. В качестве примера здесь представлено образование АТФ из креатинфосфата - соединения, которое служит в мышцах энергетическим ресурсом (см. с. 328). Формально перенос фосфатной группы с креатинфосфата на АДФ является суммарной реакцией гидролиза креатинфосфата (а) и синтеза АТФ (б).
В. Субстратное
фосфорилирование
Кроме окислительного фосфорилирования, в
промежуточном метаболизме животных только в двух реакциях неорганический
фосфат (Рi) переносится на АДФ (или соответственно ЦДФ) за счет
высокого химического потенциала. Такие процессы называют 'субстратным
фосфорилированием', поскольку они являются частью метаболического пути
('субстратной цепи'). Один из таких промежуточных этапов - образование ГТФ в
цитратном цикле (см. с. 138); вторая такая реакция, ответственная за образование
макроэргических связей, осуществляется в процессе гликолиза (см. с.
152).
На схеме представлена реакция, катализируемая глицеральдегид-3-фосфатдегидрогеназой.
Сначала SH-группа остатка цистеина молекулы фермента присоединяет карбонильную
группу глицеральдегид-3-фосфата (а). Этот промежуточный
продукт 1 окисляется НАД+ с образованием макроэргической тиолсложноэфирной
связи (б). На третьей стадии (в)
неорганический фосфат замещает тиол с образованием смешанного ангидрида 1,3-дифосфоглицерата.
В этом соединении фосфатный остаток обладает настолько высоким потенциалом,
что на следующей стадии может переноситься на АДФ (не показано, см. с. 148).