


136-137
Метаболизм. Энергетика
Дегидрогеназы кетокислот
 В промежуточном метаболизме имеются
мультиферментные комплексы, катализирующие сложную многостадийную реакцию
окислительного декарбоксилирования 2-кетокислот и переноса образующегося
ацильного остатка на кофермент А. В качестве акцептора электронов выступает
НАД+. Кроме того, в реакции участвуют тиаминдифосфат, липоамид и ФАД.
К дегидрогеназам кетокислот относятся:
В промежуточном метаболизме имеются
мультиферментные комплексы, катализирующие сложную многостадийную реакцию
окислительного декарбоксилирования 2-кетокислот и переноса образующегося
ацильного остатка на кофермент А. В качестве акцептора электронов выступает
НАД+. Кроме того, в реакции участвуют тиаминдифосфат, липоамид и ФАД.
К дегидрогеназам кетокислот относятся:
а) пируватдегидрогеназный комплекс (ПДГ,
пируват→ацетил-КоА),
б) 2-оксоглутаратдегидротеназный комплекс цитратного цикла
(ОГД, 2-оксоглутарат→сукцинил-КоА) и
в) участвующий в катаболизме разветвленных
цепей валина, лейцина и изолейцина дегидрогеназный комплекс (см. с. 402).
В
качестве примера здесь рассмотрен ПДГ-комплекс.
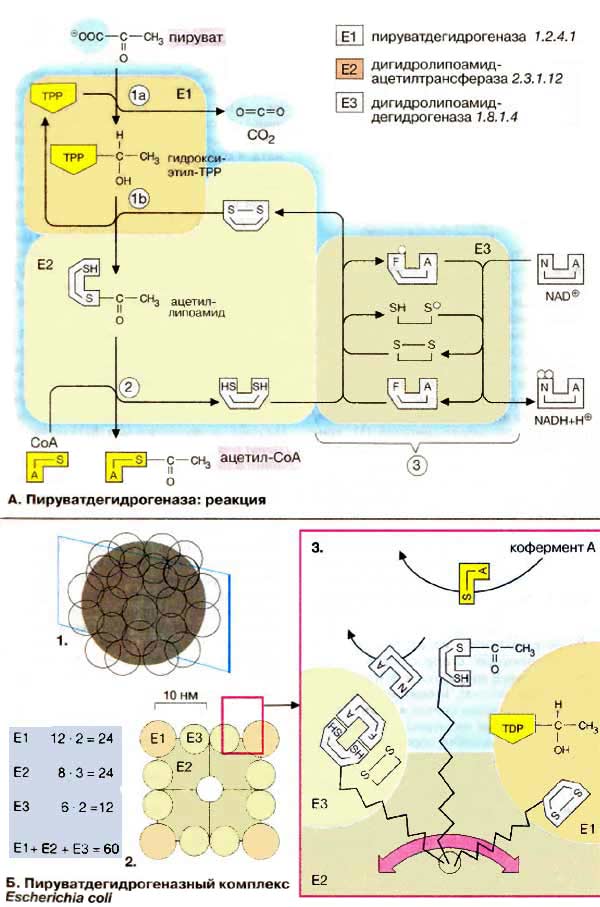
А. Пируватдегидрогеназа: реакция
В пируватдегидрогеназной реакции участвуют три различных фермента
[1-3]. Пируватдегидрогеназа (Е1) катализирует декарбоксилирование
пирувата, перенос образованного гидроксиэтильного остатка на тиаминдифосфат
(TPP, 1а), а также окисление гидроксиэтильной группы
с образованием ацетильного остатка. Этот остаток и полученные восстановительные
эквиваленты переносятся на липоамид (1б). Следующий
фермент, дигидролипоамидацетилтрансфераза (Е2) переносит ацетильный остаток
с липоамида на кофермент А (2), при этом
липоамид восстанавливается до дигидролипоамида. Последний снова окисляется до
липоамида третьим ферментом, дигидролипоамиддегидрогеназой (Е3)
с образованием НАДН + Н+ (NADH + Н+) (3).
Электроны переносятся на растворимый НАД+ через ФАД и каталитически
активный дисульфидный мостик субъединицы Е3.
Пять разных коферментов этой
реакции различными способами ассоциированы с белковыми компонентами ферментов.
Тиаминдифосфат нековалентно связан на Е1, Липоамид ковалентно связан с остатком
лизина Е2, а ФАД прочно ассоциирован в виде простетической группы на Е3.
НАД+ (NAD+) и кофермент А взаимодействуют с комплексом в
виде растворимых коферментов.
Б. Пируватдегидрогеназный комплекс
Escherichia coli
Пируватдегидрогеназный комплекс
(ПДГ-комплекс) бактерии Escherichia coli достаточно подробно исследован.
Он имеет молекулярную массу от 5,3 · 106 Да и диаметр больше 30 нм,
т. е. ПДГ-комплекс крупнее, чем рибосома. Комплекс состоит из 60 полипептидов
(1,2): 24 молекулы Е2 (8 тримеров) образуют ядро комплекса кубической формы.
Каждая из 6 граней этого куба занята димерами (возможно, тетрамерами)
компонентов Е3, в то время как на 12 ребрах куба лежат димеры молекул Е1. Другие
дегидрогеназы кетокислот построены аналогично, но могут отличаться числом
субъединиц и молекулярной массой.
Пространственная организация составных
частей комплекса очень важна для катализа. Кофермент, липоевая кислота, очень
подвижен благодаря образованию связи с лизиновым остатком фермента Е2.
"Ручка" липоамида длиной примерно 1,4 нм в процессе катализа перемещается
между E1 и Е3 (3). Липоамид таким способом может взаимодействовать как со
связанным в Е1 тиаминдифосфатом, так и с растворимым коферментом А и
акцептирующим электроны ФАД в Е3. Белковый домен ацетилтрансферазы, который
связывает липоевую кислоту, очень гибок. Это дополнительно повышает дальность
действия липоамидной 'ручки'.
Регуляция ПДГ-комплекса организма животных обсуждается
на с. 152.