


166-167
Метаболизм липидов
Деградация жирных кислот: β-окисление
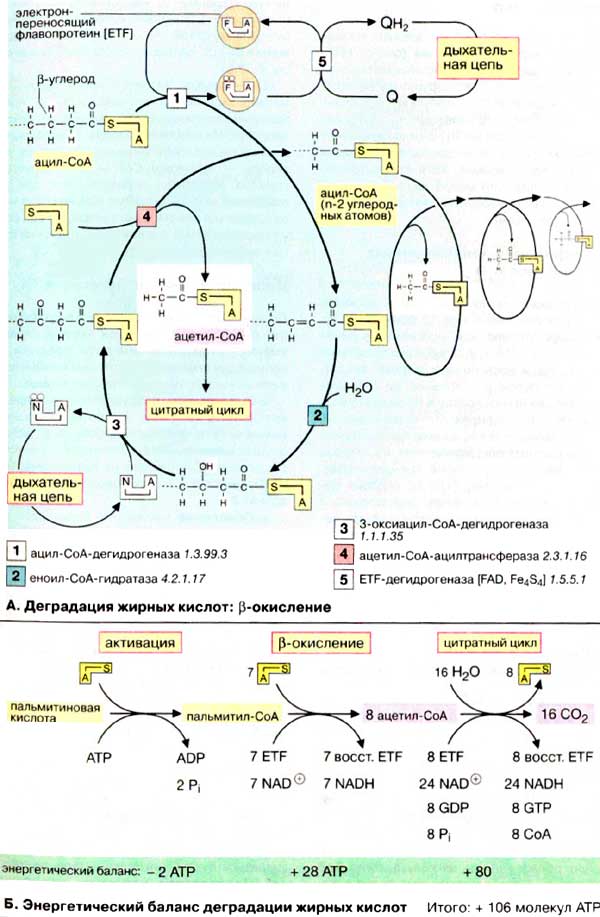 А. Деградация жирных кислот:
β-окисление
А. Деградация жирных кислот:
β-окисление
После попадания в клетки жирные кислоты
активируются путем образования ацил-КоА Для этого нужны две богатые энергией
ангидридные связи АТФ (см. с. 112). В матрикс митохондрий активированные жирные
кислоты попадают в виде ацилкарнитина, который является трансмембранным
переносчиком (см. с. 214).
Деградация жирных кислот происходит в
митохондриальном матриксе путем окислительного цикла реакций, при котором
последовательно отщепляются С2-звенья в виде ацетил-КоА
(активированной уксусной кислоты). Последовательное отщепление
ацетильных групп начинается с карбоксильного конца активированных жирных кислот
каждый раз между С-2 (α-атомом) и С-3 (β-атомом). Поэтому цикл реакций
деградации называется β-окислением. Пространственно и функционально
β-окисление тесно связано с цитратным циклом (см. с. 140) и дыхательной цепью
(см. с. 142).
Первая стадия β-окисления - дегидрирование активированной
жирной кислоты (ацил-КоА) с образованием β-ненасыщенной жирной кислоты
с двойной связью в транс-конфигурации (реакция [1]:
дегидрирование). При этом оба атома водорода с электронами переносятся
от фермента [1] на электронпереносящий флавопротеин
(ETF). ETF-дегидрогеназа (5) переносит
восстановительные эквиваленты на убихинон (кофермент Q), который является составной
частью дыхательной цепи (см. рис. 143). Вторая стадия
деградации жирной кислоты состоит в присоединении молекулы воды к двойной
связи ненасыщенной жирной кислоты (реакция [2]:
гидратирование). На третьей стадии происходит окисление гидроксильной
группы при С-3 в карбонильную группу (реакция [3]:
дегидрирование). Акцептором для восстановительных эквивалентов является
НАД+ который передает их в дыхательную цепь. На четвертой
стадии активированная β-кетокислота расщепляется ацилтрансферазой (β-кетотиолазой)
в присутствии кофермента А (реакция [4]: тиолитическое
расщепление). Продуктами реакции являются ацетил-КоА и активированная жирная
кислота, углеродная цепь которой короче на два углеродных атома по сравнению
с длиной цепи исходной жирной кислоты.
Для полной деградации длинноцепочечной
жирной кислоты цикл должен многократно повторяться; например, для стеарил-КоА
(18:0) необходимы восемь циклов. Образующийся ацетил-КоА может переноситься на
оксалоацетат с образованием цитрата, промежуточного метаболита цитратного
цикла (см. с. 140). При избытке ацетил-КоА в печени образуются кетоновые
тела (см. с. 304).
Б. Энергетический баланс деградации
жирных кислот
Для расчета энергетического баланса деградации жирной кислоты
в качестве примера рассмотрим молекулу пальмитиновой кислоты (16:0),
которая окисляется полностью до 16 молекул СО2. На первой стадии
жирная кислота активируется, потребляя две богатые энергией связи [АТФ (АТР)],
с образованием пальмитоил-СоА состоящего из восьми C2-звеньев.
Затем протекают семь циклов β-окисления. При этом образуются 7 молекул
восстановленной формы флавопротеина (ETF) и 7 молекул НАДН + Н+.
Оба соединения включаются в дыхательную цепь; окисление ETF через убихинон дает
в итоге 1,5 молекулы АТФ, а НАДН + Н+ - 2,5 молекулы (см. рис. 143).
Таким образом, β-окисление одного пальмитоильного остатка дает 28 молекул
(7 х 4) АТФ. Окисление каждой молекулы ацетил-КоА приводит к образованию
10 молекул АТФ, что означает получение еще 80 молекул (8 x 10) АТФ. Из 28 +
80 молекул АТФ следует вычесть две молекулы АТФ, израсходованные при активации
пальмитиновой кислоты (см. выше). Итак, при утилизации одной молекулы пальмитиновой
кислоты синтезируются 106 молекул АТФ, что соответствует свободной энергии 3300
кДж/моль (106 х 30,5 кДж/моль АТФ). Выигрыш в энергии при деградации жирных
кислот существенно выше по сравнению с распадом углеводов (32 молекулы АТФ на
1 молекулу глюкозы) и белков даже с учетом больших размеров молекул. Поэтому
жиры представляют собой очень выгодную форму сохранения энергии.