


90-91
Биомолекулы. Нуклеиновые кислоты
Дезоксирибонуклеиновые кислоты
 Биологическая функция нуклеиновых кислот
(см. с. 234) основана главным образом на свойстве оснований образовывать
специфически (комплементарно) связанные пары.
Биологическая функция нуклеиновых кислот
(см. с. 234) основана главным образом на свойстве оснований образовывать
специфически (комплементарно) связанные пары.
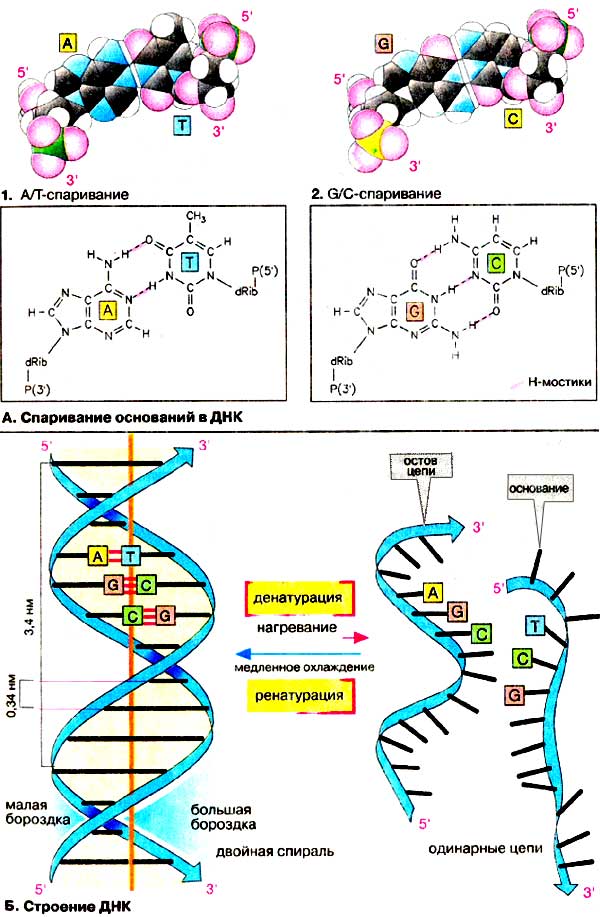
А. Спаривание оснований в ДНК
Первым свидетельством существования таких структур пocлyжил
тот факт, что в каждом типе ДНК (DNA) содержится примерно одинаковое количество
аденина и тимина. То же самое относится к гуанину и цитозину. Напротив, соотношение
(аденин+тимин) / (гуанин+цитозин) в различных организмах варьирует. Предложенная
в 1953 г. модель структуры ДНК позволила объяснить причину таких соотношений:
интактная ДНК состоит из двух полидезоксинуклеотидных цепей. Каждое
основание одной цепи связано с комплементарным ему основанием другой
цепи водородными мостиками. При этом аденин комплементарен тимину, гуанин
- цитозину. Таким образом, каждая пара состоит из одного пуринового и одного
пиримидинового основания.
Комплементарность А и T, соответственно G и С, становится понятной,
если рассмотреть возможные водородные мостики между основаниями. В качестве
доноров (см. с. 14) выступают аминогруппы (аденина, цитозина,
гуанина) и NН-группы гетероциклов (тимина и гуанина). Возможными акцепторами
являются карбонильные группы (тимина, цитозина, гуанина) и атомы азота гетероциклов.
Пара A-T может образовывать два, а пара G-C даже три линейных
и поэтому особенно устойчивых мостика. Урацил, содержащийся в РНК вместо тимина,
ведет себя при спаривании основании подобно тимину.
Б. Структура ДНК
Спаривание оснований, проиллюстрированное на
схеме А, охватывает в молекуле ДНК миллионы звеньев. Конечно, это возможно
только в том случае, если полярность обеих цепей различна, т.е. обе цепи
имеют противоположные направления (см. с. 86). Кроме
того, обе цепи должны быть закручены в виде двойной спирали. Из-за
стерических ограничений, вызванных 2'-ОН-группой остатка рибозы, РНК не мoгyт
образовывать структур, подобных двойной спирали. Поэтому РНК имеют менее
регулярную структуру по сравнению с ДНК (см. с. 88).
Преобладающая в клетке конформация ДНК (так называемая В-ДНК)
представлена схематически на схеме 1, а также на
с. 92, в виде вандерваальсовой модели. На схеме 1
дезоксирибозофосфатный остов изображен в виде ленты. Основания (здесь указаны
в виде полос) расположены внутри двойной спирали. Следовательно, эта
область ДНК неполярна.
Напротив, внешняя сторона молекулы полярна и
заряжена отрицательно за счет углеводных остатков и фосфатных групп остова. Цепи
ДНК на протяжении всего тяжа образуют два желоба, которые носят названия
'малая бороздка' и 'большая бороздка'.
Так как обе цепи связаны только
нековалентными взаимодействиями, двойная спираль при нагревании или инкубации в
щелочном растворе легко распадается на отдельные цепи
(денатурирует). При медленном охлаждении ранее неупорядоченные отдельные
цепи благодаря спариванию оснований вновь образуют двойную спираль (молекула
ренатурирует). Процессы де- и ренатурации играют важную роль в генной инженерии
(см. сс. 254, 258).
В функциональном отношении две цепи ДНК не
эквивалентны. Кодирующей цепью (матричной, смысловой) является та из них,
которая считывается в процессе транскрипции (см. с. 240). Именно эта цепь служит
матрицей для PHK. Некодирующая цепь (антисмысловая) по
последовательности подобна РНК (при условии замены T на U). Общепринято
давать структуру гена в виде последовательности некодирующей цепи
ДНК в направлении 5'→3'. Если прочитать кодоны в этом направлении, то с
помощью генетического кода (см. с. 244) можно воспроизвести аминокислотную
последовательность белка в принятом порядке, от N- к С-концу.