


192-193
Метаболизм нуклеотидов
Биосинтез нуклеотидов
 А. Синтез нуклеотидов: общие
сведения
А. Синтез нуклеотидов: общие
сведения
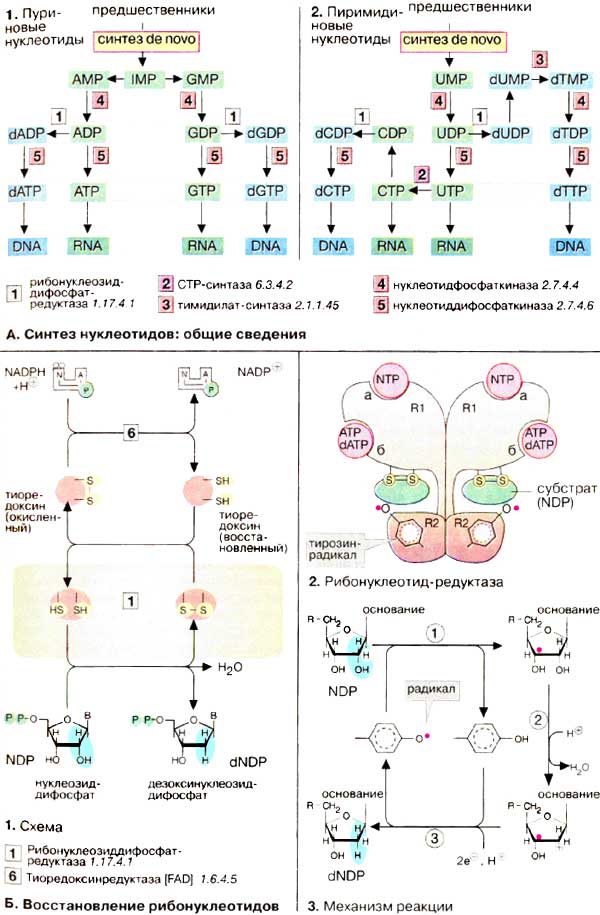
Синтез пуринов и пиримидинов de novo приводит к монофосфатам,
соответственно ИМФ (IMP] и УМФ (UMP) (см. с. 190). Из
этих двух предшественников синтезируются все другие нуклеотиды. Соответствующие
пути представлены в данном разделе. Подробности приведены на сс. 405
и 406. Синтез нуклеотидов путем повторного использовании
оснований рассмотрен на с. 188.
Синтез пуриновых нуклеотидов (1)
осуществляется из инозинмонофосфата [ИМФ (IMP)]. Его основание гипоксантин
превращается в две стадии соответственно в аденин или гуанин. Образующиеся
нуклеозидмонофосфаты АМФ (AMP) и ГМФ (GMP) переходят в дифосфаты
АДФ (ADP) и ГДФ (GDP) под действием нуклеозидфосфаткиназ и,
наконец, фосфорилируются нуклеозиддифосфаткиназами до трифосфатов АТФ
(АТР) и ГТФ (GTP). Нуклеозидтрифосфаты служат строительными блоками
для РНК (RNA) или функционируют в качестве коферментов (см. с. 110).
Преобразование рибонуклеотидов в дезоксирибонуклеотиды происходит на стации
дифосфатов и катализируется нуклеозиддифосфат-редуктазой (схема Б).
Пути биосинтеза пиримидиновых
нуклеотидов (2) сложнее, чем пути синтеза пуриновых нуклеотидов. Прежде
всего исходный УМФ (UMP) фосфорилируется до ди-, а затем трифосфата
УТФ (UTP). УТФ превращается цитидинтрифосфат-синтазой
(CTP-синтаза) в ЦТФ (СTP). Так как восстановление пиримидиновых
нуклеотидов до дезоксирибонуклеотидов происходит на стадии дифосфатов, ЦТФ
должен быть гидролизован фосфатазой до ЦДФ (CDP), после чего могут
образоваться дЦДФ (dCDP) и дЦТФ (dCTP).
Строительный блок ДНК (DNA),
дезокситимидинтрифосфат [дТТФ (dTTP)], синтезируется из УДФ (UDP) в
несколько стадий. Основание тимин, которое, по-видимому, находится только в ДНК
(см. с. 86), образуется на уровне нуклеозидмонофосфата при метилировании
дезоксиуридинмонофосфата. Отвечают за эту стадию тимидилат-синтаза и
вспомогательный фермент дигидрофолат-редуктаза, которые являются важными
мишенями для действия цитостатиков (см. с. 388).
Б. Восстановление
рибонуклеотидов
2'-Дезоксирибоза, структурный элемент
ДНК, не синтезируется в виде свободного сахара, а образуется на стадии дифосфата
при восстановлении ρибонуклеозиддифосфатов. Такое восстановление - сложный
процесс, в котором участвует несколько белков. Необходимые восстановительные
эквиваленты поставляются НАДФН (NADPH). Тем не менее, они не переносятся
непосредственно от кофермента к субстрату, а проходит прежде всего через ряд
окислительно-восстановительных реакций. На первой стадии (1)
тиоредоксинредуктаза восстанавливает с помощью связанного с ферментом
флавинадениндинуклеотида небольшой белок, тиоредоксин. При этом
дисульфидный мостик в тиоредоксине расщепляется. Образующиеся SH-группы снова
восстанавливают каталитически активный дисульфидный мостик в
нуклеозиддифосфат-редуктазе ('рибонуклеотид-редуктаза'). Свободные
SH-группы являются действенными донорами электронов для восстановления
рибонуклеотиддифосфатов.
Рибонуклеотид-редуктаза эукариот представляет собой тетрамер,
состоящий из двух R1- и R2 -субъединиц. Кроме упомянутого дисульфидного мостика,
в ферменте во время реакции образуется тирозин-радикал (2,
см. с. 20). генерирующий радикал в субстрате (3).
Последний отщепляет молекулу воды и вследствие этого переходит в радикал-катион.
При последующем восстановлении образуется остаток дезоксирибозы и регенерируется
тирозиновый радикал.
Процесс регуляции
рибонуклеотид-редуктазы имеет довольно сложный механизм. Субстратная
специфичность и активность фермента контролируются двумя аллостерическими
центрами связывания (а и б) R1-субъединицы. АТФ и дАТФ (dATP) соответственно
повышают и уменьшают активность редуктазы, связываясь с центром а. С центром б
взаимодействует другой нуклеотид, изменяющий в результате связывания субстратную
специфичность фермента.