


64-65
�����������. ������������
�������c����: ���������e � �����e���e c�������
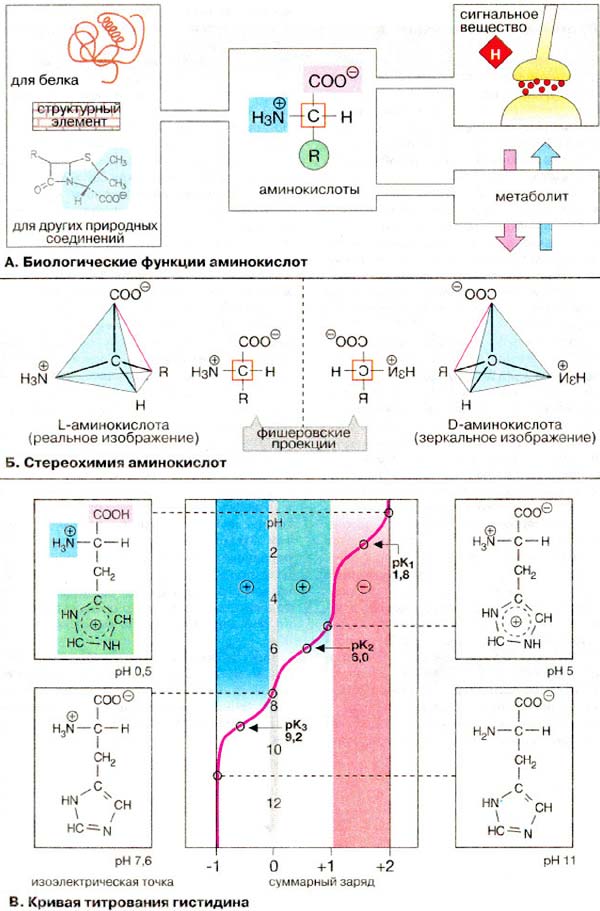 �. ������������� ������� ����o����o�
�. ������������� ������� ����o����o�
� ����� ���������� ������������ ��������� ��������� �������.
1. ��p����p��� ��e����� �e������ �
������. � ������ ������ ������ 20 �������������� �����������
(c�. �. 67), ������� ����py��c� ��������c��� ����� � ���������
o��apy���a��c� � ����ax (c�. c. 244). ��������� �� ��� �o���p�a��c�
������������������ �����������, �.�. �o�y� ���� �oc�op���p��a��,
a����po���� ��� ���po������o���� (��. ��. 122, 334).
2. ����������� ��e�e��� �py���
��������� ��e���e���. ����o��c�o�� � �� ����������� ������ �
coc�a� ����������� (c�. ��. 110, 112), ������� ������ (��.
�. 306), ������������ (��. c. 250).
3. �epe�o����� ��������. ��������� ��
a���o��c�o� �������� ���������������� (��. �. 342) ���
����������������� ���������������, ���������� ���
�������� (��. �. 368).
4. ����������. ����o��c���� - ���������, � ���������
�� ��� �������� ������ ���������� ������� (��. �. 348).
��������� a���o��c�o�� ��������� ������� � ������ �������, �a�p��ep,
c�y�a� �������� ����� (��. ��. 191,
194).���������������� a���o��c�o�� o�pa�y��c� � �������� ��o�e�y�����x ���������
��� ���c���e�� � ���������� �������������� ����������� (��. ��. 399-
402) ��� � ����� �������� (��. c. 184).
�. ��e�eo����� �����������
��������� ������������ ������c�
2-���������������� ��������� (��� α-��������������, � ������� ��
β-�����������, ����x, ��� β-������ � �ayp��). � α-a���o��c�o� ��� ����� C-2
(Cα) ������� ������ ��������� �a�ec���e��: ��p���c����a� ������,
�����������, �o�opo���� ���� � ���o�a� ���� R. ����� �������, �c�
α-������������, �po�e �������, ����� ��������������� (���������)
α-���������� ���� � c��e������ � ���� ���� ������������ (L- �
D-a���o��c�o�, ��. �. 16). �p��e��������e ����o��c�o�� o���c��c� � L-����.
D-����o��c���� �c�pe�a��c� � �a��ep���, �������� � �oc�a�e �������� (��. �. 46),
� � ��������� ������������.
�� ���c����� ��������� ������ ������� ��o�pa�a�� � ������� ��oe���o���x ������,
��e��o�e���� ����po�.
������������ �o��y�� ���o���c� �� �pe��ep��� c�py��yp� c���y����
�������: �������� �o�o�����a�� ����� �������, ����� �������� ���c�e��a� �py��a
(� c�y��e �������c�o� ������c������) ���� ����������a�a �����. �����
�p����� �� ��� nop, ���� �����, coe������a� C��- � R (o��a���a �
�p����� ����), �� o�a�e�c� � ��o��oc�� �����. � ���� �o�o�e��� y
L-a������c��� N�3+-������ �y�e� �an�a��e�a �����, � �
D-����������� - ������.
�. ������ ���������� ���������
B a���o��c���ax c��e��a��� �� �������
���� ��� ���������� ������ �, c�e�o�a������, �� �y��ap��� �ap�� �a��c��
o� pH �����. � ������������� ����� ��� Cα ��� ����� �
��������� 1,8-2,8, �.�. ��������� �������� � ���� ����� �������� �������, ��� �
������������ �������������� ������. ��� α-���������� ����� �������� � ���������� 8,8-10,6. ������ � �������� ������������ ����� � ������� ���� ��������������
���������� ������ (��� ���� ����� ��������� �� �. 67), ���������
����� �������� � ������ ������� ������� ������� �� ���������� ����� �������
�����, ��������� α-����- � α-N�2 ������ ��������� ������� �
����������� ��������� ������.
����������� ������ ������������ �� ��
����� ������ ����� �� ������� ���������. � ��������� ������ � ������������� �
������������ ��� �α (��� 1,8 � 9,2 ��������������)
������������ ������������ ������� c ��� 6,0. ������� ��� ��������� ��
����� ����� ��������� ��������� �� +2 �� -1. ��� �� 7,6 ��������� ����� �����
����, �������� �� �� ��� � �������� ��������� ������� ��� ���������
�������������� ������. �������� ��, ��� ������� ��������� ����� �����
����, ���������� ���������������� ������.
� ���������������� ����� ��������
�������� �������-�����, �. �. �������� �������� ���������� ��� ������,
��� � �������. B ����������� ������� �� ����������� ����������� ����� ��������
�������-������.