


108-109
Метаболизм. Ферменты
Окислительно-восстановительные коферменты
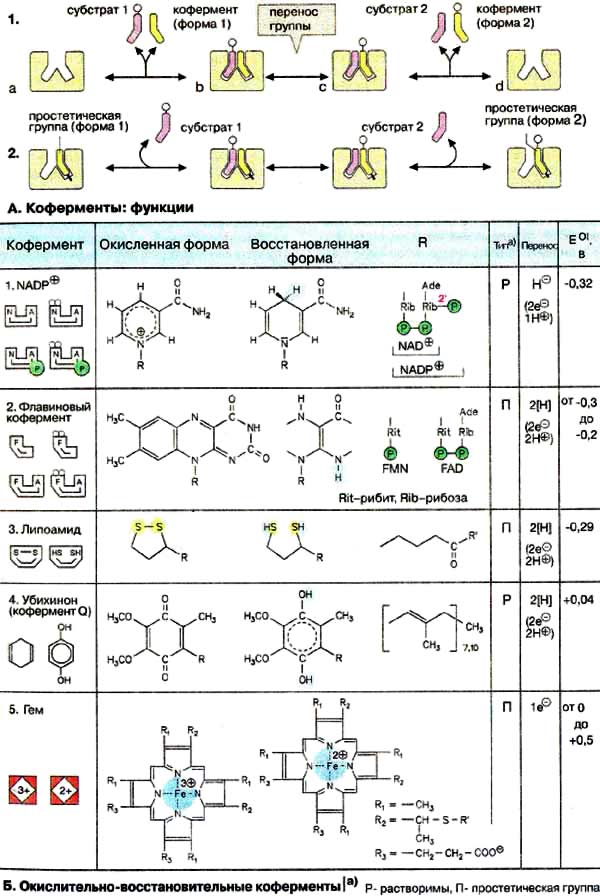 А. Коферменты: функции
А. Коферменты: функции
Многие ферментативные реакции включают
перенос электронов или групп атомов с одного субстрата на другой. В таких
реакциях всегда принимают участие вспомогательные соединения
(коферменты), которые выполняют функцию промежуточных переносчиков атомов
или функциональных групп. Так как эти вещества каталитически не активны,
правильнее было бы их называть косубстратами. Ферменты обычно
высокоспецифичны к своим субстратам (см. с. 94), коферменты же взаимодействуют
со многими ферментами, обладающими различной субстратной
специфичностью.
По способам взаимодействия с ферментом различают растворимые
коферменты и простатические группы. Растворимый кофермент (1)
присоединяется во время реакции к молекуле фермента подобно субстрату,
химически изменяется и затем снова освобождается. Первоначальная форма
кофермента регенерируется во второй, независимой реакции. Простетической
группой (2) называется кофермент, который прочно
связан с ферментом и во время реакции его не покидает. Группа, связавшаяся с
коферментом, далее переносится на следующий субстрат или другую молекулу кофермента
(на схеме 2 не показано).
Б. Окислительно-восстановительные
коферменты
Все оксидоредуктазы (см. с. 94) нуждаются в
коферменте. Наиболее важные коферменты представлены на схеме. Они могут
действовать в растворимой форме (Р) или в виде простетической группы (П).
Окислительно-восстановительные реакции, наряду с переносом электронов, часто
включают перенос одного или двух протонов. Поэтому обычно принято говорить о
переносе восстановительных эквивалентов. Стандартный потенциал
Eo' простетической группы (см. с. 24) может значительно отличаться в
зависимости от окружения в молекуле фермента.
Пиридиннуклеотиды НАД+
(NAD+) и НАДФ+ (NADP+) (1) широко
распространены как коферменты дегидрогеназ. Они
переносят гидрид-ион (2е- и 1 H+, см. с. 102) и
действуют всегда в растворимой форме. НАД+ передает
восстановительный эквивалент из катаболического пути вдыхательную цепь и тем
самым участвует в энергетическом обмене. HАДФ+, напротив, является
самым важным восстановителем при биосинтезе (см. с. 114).
Флавиновые коферменты ФМН (FMN) и ФАД (FAD) (2,
см. с. 86) найдены в дегидрогеназах, оксидазах и
монооксигеназах. Обычно оба соединения ковалентно связаны с ферментами.
Активной группой обоих коферментов является флавин (изоаллоксазин), имеющий
сопряженную систему из трех колец, которая может при восстановлении принимать
два электрона и два протона. В ФМН к флавину присоединен фосфорилированный полиол
рибит. ФАД состоит из ФМН, связанного с АМФ. Оба соединения являются
функционально близкими коферментами.
В липоевой кислоте (3) функцию
окислительно-восстановительного центра выполняет внутримолекулярный
дисульфидный мостик. Активная липоевая кислота ковалентно связана
с остатком лизина (R') молекулы фермента. Липоевая кислота прежде всего
участвует в окислительном декарбоксилировании 2-кетокислот (см. с. 136).
Дисульфидный мостик также содержится в пептидном коферменте глутатионе
(см. с. 278).
Функция убихинона (кофермента Q , 4)
как переносчика восстановительного эквивалента в дыхательной цепи будет рассмотрена
на с. 142. При восстановлении хинон превращается в ароматический
гидрохинон (убихинол). Похожие системы хинон/гидрохинон принимают участие
в реакциях фотосинтеза (см. с. 132). К этому классу окислительно-восстановительных
систем принадлежат также витамины Е и К (см. с. 352).
Группа гема (5) является
окислительно-восстановительным кофактором в дыхательной цепи (см. с. 144),
фотосинтезе (см. с. 130), а также в монооксигеназах
(см. с. 310) и пероксидазах. В отличие от гемоглобина
в этих случаях ион железа меняет валентность. На рисунке показан гем в цитохроме
с, ковалентно связанный с двумя остатками цистеина (R2) белка.